
100���±�ţ�1004-0609(2010)02-0346-08
��ϸ��¾�������ͭ��������������Ⱥ�������
�� ��1������ѩ1���� ��1, 2��������1, 2����ѧ��1, 2
(1. ���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ����ɳ 410083��
2. ���ϴ�ѧ ����ұ��������ص�ʵ���ң���ɳ 410083)
ժ Ҫ���о�3�꼫�����ȹž�(������Ҷ����Sulfolobus metallicus JCM 9184��ɪ�������������Metallosphaera sedula JCM 9185�����������Acidianus manzaensis YN25)�ڲ�ͬ��ʼpHֵ�Ͳ�ͬ�¶������¶Ի�ͭ��Ļ�Ͻ��������Խ�������л�Ͼ�Ⱥ��Ķ�̬������з������������������ʼpH 1.5ʱ��ͭ���������Ը�������ʼpH 2.5ʱ��ͭ�����ʣ���65 �������µ�ͭ�����ʸ���75 ��ʱ��ͭ�����ʡ����������Գ��ȶ�̬��(RFLP)����65 �桢��ʼpH 1.5�����µ�����Ⱥ�����棬�����ʾ���ڻ�ͭ��Ľ���ǰ��Sulfolobus metallicus��ռ�����Ƶľ��֣���������Acidianus manzaensis�ı�����������������ȡ��Sulfolobus metallicus��Ϊ�����֡�
�ؼ��ʣ�
�����������ͭ����������������������Ⱥ����������
��ͼ����ţ�Q 939���� ���ױ�ʶ�룺A
Bioleaching of chalcopyrite concentrate using
mixed thermophilic culture and succession of
microbial community during leaching process
Liu Kai1, Diao Meng-xue1, Yang Yu1, 2, Qin Wen-qing1, 2, Wu Xue-ling1, 2
(1. School of Minerals Processing and Bioengineering, Changsha 410083, China;
2. Key Laboratory of Biometallurgy, Ministry of Education, Central South University, Changsha 410083, China)
Abstract: A mixed thermophilic culture consisting of Sulfolobus metallicus JCM 9184, Metallosphaera sedula JCM 9185 and Acidianus manzaensis YN25 was used to study the combined effect of pH value and temperature in bioleaching of a chalcopyrite concentrate. The results show that the copper extraction at initial pH 1.5 is higher than that at intial pH 2.5. The copper extraction at 75 �� is lower than that at 65 ��, but the lower copper extraction at 75 �� is not due to the inhibition of the high temperature. The succession of the microbial community during the leaching process was analysed by restriction fragment length polymorphism (RFLP). The analysis shows that Sulfolobus metallicus is dominant in the microbial community at the early stage of leaching, whereas Acidianus manzaensis thrives at the latter stage. This phenomenon might be caused by the change of ferrous ion concentration during leaching process.
Key words: bioleaching; chalcopyrite concentrate; extremely thermophilic microorganisms; microbial community; succession
��ͭ������Ȼ������Ҫ��ͭ�����������������ṹ���������ʺ������ʻ�ͭ������ѳ�Ϊͭ��ҵ��ע���ȵ㡣�봫ͳ������ȣ�����������й��ռ��ɱ��ͺͶԻ����Ѻõ��ŵ㡣���������ͭ�����漰�Ļ�ѧ��Ӧʽ����[1]�����У��������ķ�Ӧ�з�Ӧʽ(3)�ͷ�Ӧʽ(4)��
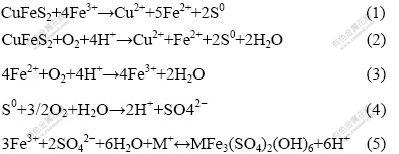
ͨ���Ա����¾����е����Ⱦ��ͼ������ȹž��ֱ������ͭ���ʵ������֪������Ч�����¶ȴ��³������[2?3]���������¾�����ʱ(35 ��)�����ڽ��������п��������γ�Ԫ�����м�����ͻƼ����������ʶ�ʹ����ۻ���ͭ�Ľ������ʺͽ����ʽ� ��[4?7]��ͨ����30%����[8]�����ڽϸߵ��¶���(68 ��)�������м���������ܽ�[7]��ͭ�����ʻ����ߣ�һ��ɴ�90%����[8]��STOTT��[9]����Sulfolobus metallicus, Acidianus brierley ��Metallosphaera sedula�ȼ������Ⱦ�������ͭ��ʱ������˽Ϻõ�ͭ����Ч����BHP Billiton��˾������BioCop?�����ɹ���Ӧ�ü����������ﴦ����ͭ��ʵ���˰빤ҵ����[10]����һЩ������Ķѽ��У������������ķ��ȷ�Ӧ������ھֲ����¶Ȼ����ߵ�70~80 �棬���������������¾����е����Ⱦ�������������������������������ȴ�����������������������������ˣ����ø��¾����������ͭ���������Ч�ʵ���Ч�ֶκ�;����
��ͭ�����������������IJ��������ģ���˽�����ϵ�е�pHֵ��ͭ�Ľ���������Ҫ��Ӱ�졣�¶�ͬ���Ի�ѧ��Ӧ���ʲ����ܴ�Ӱ�죬�ڻ�ͭ����������������¶ȿ������ͭ�Ľ������ʣ����¶ȹ����ֻ����ƽ������������ʹ�ý������������������ķ��ȷ�ӦҲ��ʹ������ϵ���¶ȷ����仯���Ӷ�Ӱ�����Ч�ʲ����������Ⱥ��ṹ�����仯����ˣ��˽��ڿ�����������л�Ͻ��������Ⱥ��ṹ������������л����ľ��꣬����Ϊ�Ż��Ϳ��ƽ��������ṩ���õ���Ϣ��Ŀǰ�����в��ٹ�������ұ���������������Ժ�Ⱥ����о���������������������������е�����������Ϊ�о������ڽ�������м�����������Ⱥ��ṹ�仯�ı���������������Ҿͻ�ͭ����ԣ�����������һ�����Ż��Ľ���������ϻ���Ҫ��һ�����о���̽�� [11]����ˣ�������������3�꼫�����������ڲ�ͬ�¶ȡ���ͬpH�����Ի�ͭ����л�Ͻ������������������еĹž�Ⱥ�䣬�˽���¾��ڻ�ͭ����������е����á�����Ч����Ӱ�����غ�Ⱥ��仯��Ϊ��һ���Ŀ�����Ӧ�ü����������������ͭ���ṩ���ݡ�
1 ʵ��
1.1 ʵ�����
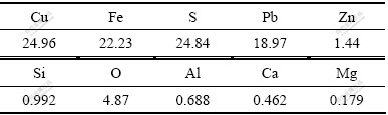
ʵ�����õĻ�ͭ��������ұ��������ص�ʵ�����ṩ�������������顢��ĥ��ɸ�֣�����С��74 ��m������Ԫ�ط�������ѧ�����������1��2��
��1 ������ѧԪ�ط���
Table 1 Element analysis of sample (mass fraction, %)
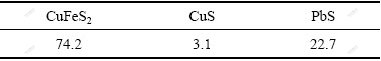
��2 ������ѧ�������
Table 2 Component analysis of sample (mass fraction, %)
1.2 ���ּ���������
��ʵ�����õ��ļ������ȹž�������������Ҷ��(Sulfolobus metallicus JCM 9184)��ɪ�����������(Metallosphaera sedula JCM 9185)���������(Acidianus manzaensis YN25)��ǰ���������ձ����ﱣ������(JCM)����һ���ɱ�ʵ���Ҵ������ڳ����Ȫ�з���õ���3�������9K����������[12]�ӽ�ĸ�����࣬�Ե��������������Ϊ��Դ���ʴ����������䷽���£�(NH4)2SO4 3 g/L��KCl 0.1 g/L��K2HPO4 0.5 g/L��MgSO4��7H2O 0.5 g/L��Ca(NO3)2 0.01 g/L����ĸ������ 0.2 g/L��S0 10 g/L����FeSO4��7H2O 30 g/L����10 mol/L�����������������pHֵ��
1.3 ��ͭ�����ʵ��
����ʵ�����500 mL����ƿ��Ϊ������ƿ��װ��200 mL���������������Ũ��4%��������������ڵ����ȹž��������ֵ���(�ֱ�Ϊ1.6��107��/ mL)��������ƿ��, ��ת��Ϊ150 r/min�ĸ���ˮԡҡ������������ÿ������������3��ƽ��ʵ��, �������հ��ա�ÿ������ȡ���ⶨ����Һ��pHֵ��������ԭ��λ����Ũ�ȡ�Cu2+��Fe2+������Ũ�ȣ�ȡ��ǰ������ˮ������������ˮ�֡�
1.4 ������ѧ����
��ԭ�����չ��ײⶨ����Һ��Cu2+��������Ũ�ȡ����ظ���صζ����ⶨ��Һ�е�Fe2+Ũ�ȡ���pHS?3C��ȼƲ���������ϵ��pHֵ���øʹ��αȵ缫�ͽ������缫�ⶨ������ϵ��������ԭ��λ���ڹ�ѧ��������Ѫ�������ⶨ����Һ������ϸ������������������X�������似����������ֵ�Ŀ�����ɡ�
1.5 �ž���DNA��ȡ��16S rDNA����
�ڽ��������У��Բ�ͬʱ��ȡ����Һ��Ʒ�������˾��ϸ��DNA��ȡ�Լ�����ȡ���йž�����DNA��16S rDNA�����������ϵ���£�33 ��L��ˮ��10��PCR buffer 5 ��L��dNTPs 5 ��L(10 mmol/L)������4F[13]��1492R[14]��2 ��L(5 pmol/��L)��Taq DNA�ۺ�ø1 ��L���ž���DNA 2 ��L���ܹ�50 ��L�������������£�94 �棬5 min��94 �棬45 s��55 �棬45 s��72 �棬1 min(��30��ѭ��)��72 �棬10 min��
1.6 16S rDNA�Ŀ�¡�������Գ��ȶ�̬�Է���(RFLP)
16S rDNA���������������pGEM-T�������Ӻ�ת��˾�DH5������̬ϸ���У�����16S rDNA�����Ŀ⡣���Ŀ�����ѡ���Կ�¡���о���PCR�������Գ��ȶ�̬�Է���������������[15]�����������Գ��ȶ�̬�Է����������ѡ���д����ԵĿ�¡�����Ϻ��������\���������ι�˾���Ը����в���Blast������GenBank��������������(http://www.ncbi. nlm.nih.gov/blast/)����ȡ���ж�Ӧ������Ϣ��
2 ���������
2.1 pHֵ�Ի�ͭ�������Ӱ��
Ŀǰ���ڽ�����ϵ��pHֵ�Ի�ͭ������ʵ�Ӱ�����Ų�ͬ�Ĺ۵㣬 VILC?EZ��[1]��Ϊ�ϵ͵�pHֵ(1.0)�ܹ��ٽ�ͭ�Ľ�������C?RDOBA��[16]����Ϊ�ϸߵ�pHֵ(2.0)������ͭ�Ľ�������Ԥʵ��Ľ��������3���������/���������е�����pHֵ�ֱ�Ϊ1.5��2.5�������¶ȷ�ΧΪ65~75 �档Ϊ�˽�һ���о�pHֵ�Ի�ͭ�������Ӱ�죬���о���65 ������������������ͬ����ʼpHֵ��1.5��2.5���Բ�ͬpH�����½���Һ�е�Cu2+Ũ�Ƚ��вⶨ��ͼ1��ʾΪ��ͬ��ʼpHֵ�����½���Һ��Cu2+Ũ�ȱ仯���ߡ���ͼ1���Կ���������ʼpHΪ1.5ʱ��������Cu2+Ũ��Ҫ���Ը�����ʼpHΪ2.5ʱ��������Cu2+Ũ�ȡ���Ϊ��24 d�Ľ��������ڣ�ǰ�ߵ�ͭ�����ʴﵽ90.8%�������ߵ�ͭ������ֻ��56.9%��
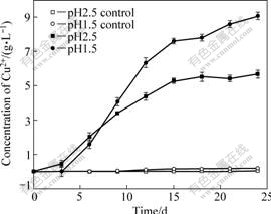
ͼ1 ��ͬ��ʼpHֵ�����½���Һ��Cu2+Ũ�ȱ仯����
Fig.1 Changing curves of Cu2+ concentration in solution at different initial pH values
��ͬpH�����½���Һ��������Fe3+��Ũ�ȱ仯��ͼ2��ʾ����ͼ2�п��Կ������ڲ�ͬ�����£�������Ũ�ȱ仯���ƶ��Ƚ����ƣ����ڽ������������ܿ죬����һ��������ֿ��ٵ��½�������ʼpHΪ2.5�����£�����Ũ���ڽ����ĵ�6��ﵽ��ߵ�(1.52 g/L)��Ȼ��Ѹ�ٵؽ��ͣ�����������Ũ�Ȳ���0.01 g/L������ʼpHΪ1.5�����£�����Ũ���ڽ����ĵ�9��ﵽ��ߵ�3.09 g/L��Ȼ��Ѹ�ٵؽ�����1.19 g/L��
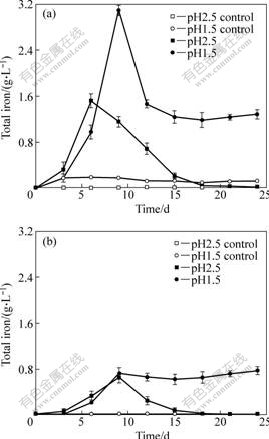
ͼ2 ��ͬ��ʼpHֵ�����½���Һ��������Fe3+Ũ�ȱ仯����
Fig.2 Changing curves of total iron (a) and Fe3+ (b) concentrations in solution at different initial pH values
����Ũ�ȵ��½���Ҫ���γ��˴�����Fe3+�������ڽ����ĺ��ڿ��Թ۲쵽����ɫ��һ��ʼ�ĺ�ɫ��������ɫ�����ҶԽ�������X�����������Ҳ֤ʵ�˴����Ƽ����������ɡ����ɵĻƼ����������ڿ�������γɶۻ��㣬�赲�����Ӫ�����ʡ������������������������Ӵ�������谭�˻�ͭ����ܽ�[17?18]����ʹ����ʼpH 1.5�������£��Ƽ��������γ���Ȼ�����ԡ�����������ʼpH 2.5ʱ��ʵ������ȣ�pHֵ�ϵ͵Ľ���Һ��ϵ�ܼ����Ƽ����������ɡ�
����Һ��pHֵ�仯�;�Ũ�ȱ仯��ͼ3��ʾ����ͼ3(a)���Կ���������ʼpH2.5�����£��ڽ�����ǰ9�죬pHֵ�����ش�2.5�½���1.5����9���Ժ�pHֵ��ʼ���������յ�1.65������Ҫ����Ϊ���������������ᵼ��pHֵ�½�������ͭ��Ļ�ѧ������һ������Ĺ���[1, 19]�������������������������ĵ���ʱ����ϵ��pHֵ�����л�����������ʼpH 1.5ʱ������������������ij���������Һ��pHֵ���½�������������ʱ�ﵽ1.16��
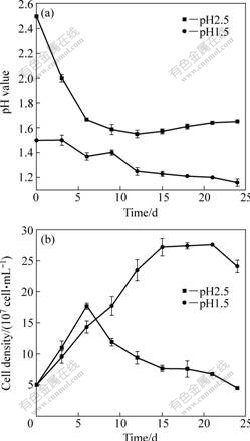
ͼ3 ��ͬ��ʼpH�����½���Һ��pHֵ�;�Ũ�ȵı仯����
Fig.3 Changing curves of pH value (a) and cells density (b) in solution at different initial pH values
2.2 �¶ȶԻ�ͭ�������Ӱ��
�Ա�������ͬpHֵ�Ի�ͭ�������Ӱ���֪�����¶�Ϊ65 �������£���ʼpHΪ1.5ʱ��ͭ�����ʸ�����ʼpHΪ2.5ʱ��ͭ�����ʡ�Ԥʵ����������3��ž��������¶���65~75 ��֮�䡣��ˣ��ڽ����ʽϸߵ�pH 1.5�����£����о��������¶ȷ�Χ�����������¶�(65��75 ��)���Ƚ��¶ȶԼ������ȹž��Ի�ͭ�������Ӱ�졣��ѧ��Ӧ����ѧ���о���������ͨ������£����ŷ�Ӧ�¶ȵ����ߣ���ѧ��Ӧ������Ҳ����Ӧ����ߣ���������¶��ܹ����ٽ��������и����ڿ�������Ԫ����������м������������Ӷ����ͭ������[20]�����ǣ����о����������75 ��������ͭ�Ľ�����Ϊ87.7%����������65 �������µ�ͭ�����ʸ�(90.8��)(��ͼ4)����ͼ4(b)���Կ�����75 �������µľ���Ũ�ȱ�65 �������µľ���Ũ�ȸ��ߣ�����������Ҳ���죬��˵���ϸߵ��¶Ȳ�û�����ƽ��������������ͭ�����ʽϵͲ��ǽϸ��¶�ֱ��Ӱ��Ľ����
`
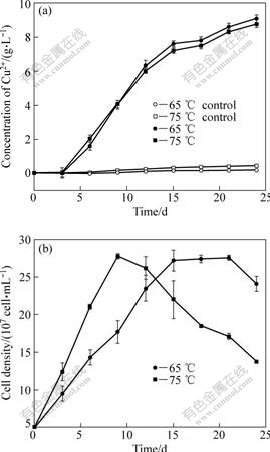
ͼ4 ��ͬ�¶������½���Һ��Cu2+Ũ�Ⱥ;�Ũ�ȵı仯����
Fig.4 Changing curves of Cu2+ concentration (a) and cells density (b) in solution at different temperatures
ͼ5��ʾΪ��65��75 �������½���Һ��������Fe3+Ũ�ȵı仯���ߡ��������ױ����������¶ȸ����������ij���[1, 21]����ͼ5�п��Է��֣�ͬ��pH 1.5�����£�75 �����ϵ������Ũ���½�������65 �����ϵ�еĴԱ�����������Fe3+Ũ�ȣ��Լ����ǵIJ�ֵ(Fe2+Ũ��)�ı仯��֪���ڽ�������65 �����ϵ�е���������Fe3+��ʽ���ڣ���75 �����ϵ�еĴ���������Fe2+��ʽ���ڡ�
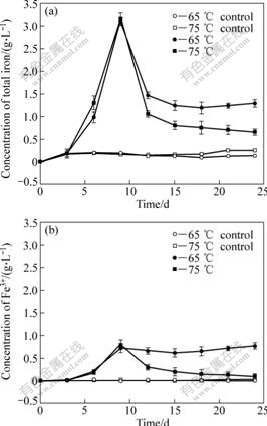
ͼ5 ��ͬ�¶��½���Һ��������Fe3+Ũ�ȱ仯����
Fig.5 Changing curves of total iron (a) and ferric ion (b) concentration in solution at different temperatures
ͬ�������½���Һ��������ԭ��λ��pHֵ�ı仯��ͼ6��ʾ����65 �������£�������ϵ��������ԭ��λ���մﵽ560 mV������75 �������£�������ϵ��������ԭ��λһֱ����400 mV��������ϵ�е��� ����ԭ��λ��Ҫ����Һ�е�Fe3+��Fe2+��Ũ�ȱ�ֵ�� ��[22]����65 �������£�����������Һ�е������Ӵ���Fe3+����75 ����������Һ�е������Ӵ���Fe2+����ϲ�ͬ�����µ�ͭ�����ʿ�֪����Һ��Fe3+Ũ�Ƚϸ�ʱͭ�Ľ�����Ҳ�ϸߡ�ͬʱ��Fe2+�Ĵ��ڶԻ�ͭ��Ľ���Ҳ����Ҫ����Ϊ���ṩ���������������Դ��ת��ΪFe3+���������ױ��������[1, 23]��
��ͼ6(b)��ʾ����������65 �滹����75 �������£����������е�pHֵ�仯���������ϱȽ����ƣ������½����ơ�ֻ���ڽ����ĵ�9�죬pHֵ�����ߣ����������ڵ�9���д�����������(��ͼ5(a))���÷�Ӧ�Ǹ�����ķ�Ӧ������75 �������£��������ڵĽ���ҺpHֵ���������(1.07)�����ֵ������ʵ�������õ����������������pHֵ[24?25]������ڽ�������ϸ��Ũ�Ƚ���(��ͼ4(b))�������½���
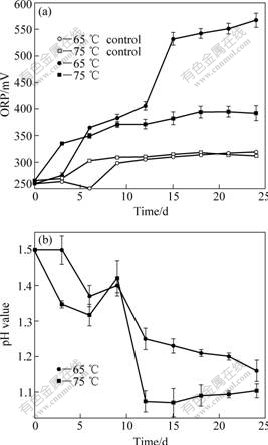
ͼ6 ��ͬ�¶��½���Һ������ԭ��λ(a)��pHֵ(b)�ı仯����
Fig.6 Changing curves of ORP (a) and pH (b) in solution at different temperatures
2.3 ������X�����������
������������X������������������ɷ֡������еĽ��������£��������ijɷֻ�����ͬ��������δ��ȫ�����Ļ�ͭ������Ǧ�ͻƼ�������ֻ�Ǹ���ֵĺ�����ͬ������ʽ(1)��(2)����ͭ����������л��е���������ɣ��������еĽ������У���û�з�����Ԫ����Ĵ��ڡ���˵��ʵ���õ��ļ������ȹž��������������ǿ�����������в�����Ԫ���������������������ᣬ��˿����Ʋ⣬��ʵ�����谭��ͭ����ȫ��������Ҫ���ز���Ԫ����Ļ��ۣ����ǻƼ������Ĵ������ڡ�����������ʼpHΪ 1.5�ҽ�������pHֵ������1.2���£�ʵ���г����ĻƼ�������Ȼ���ܽ⡣
2.4 ��������йž�Ⱥ��Ķ�̬����
�����н��������У�ѡȡ��ͭ���������ߵĽ�����ϵ(��ʼpH 1.5��65 ��)��Ϊ�о����������еĹž�Ⱥ�䶯̬���档�ֱ��ڽ����ĵ�3�졢��9�졢��15��͵�24��ӽ���Һ��ȡ����ȡ�ž�����DNA��ͨ��PCR���������ӡ�ת��������16S rDNA�Ŀ⣬�ֱ�Ӹ��Ŀ�����ȡ��70~92�����Կ�¡�ӣ���������PCR��ø�з���������Ʒ��3�ֹž��ı�����ͼ7��ʾ����ͼ7�п��Կ������ڵ�3�����Ʒ��S. metallicus��ռ�ı�����ߣ��ﵽ55%����M. sedula��A. manzaensis�ֱ��ռ27.3%��17.7%����9����Ʒ�е�������ɽṹ���3�����ƣ�S. metallicus��Ȼ�������Ƶľ��֣����ұ��������������ﵽ73.3%����M. sedula��A. manzaensis��ֱ�Ϊ18.8%��7.9%�����������е��˺���ʱ�����������Ⱥ����ɷ����˱Ƚ����Եı仯���ڵ�15�����Ʒ�У�A. manzaensis��ռ�ı�������ƷB�е�7.9%Ѹ�����ӵ�31.4%����S. metallicus��M. sedula�ı�����ֱ��½���51.5%��17.1%����һ����һֱ�����������������ڵ�24�����Ʒ�У�A. manzaensis�ı������ӵ�53.1%����S. metallicus��M. sedula�ı�����ֱ��½���37%��9.9%��
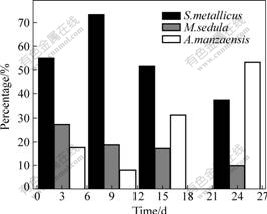
ͼ7 ��������4����Ʒ����ռ����
Fig.7 Percentages of each identified clone in 16S rDNA libraries from four samples
ͨ������Ʒ��������ĶԱȿ�֪���ڻ�ͭ��Ľ���ǰ��S. metallicus��ռ�����Ƶľ��֡������˺���A. manzaensis�ı�����������������ȡ�� S. metallicus��Ϊ�����֡���ˣ������Ʋ��ڽ��������н���Һ���������ʵĸı�Թž���Ⱥ����ɲ�����Ӱ�죬��Щ���ؿ����ǽ���Һ�еĽ�������Ũ�ȡ� pHֵ����Ϊ����������Դ��Fe2+Ũ�Ȼ�Ԫ�����Ũ�ȵȡ�
��ͼ8��ʾ���ԱȽ���Һ��Fe2+Ũ�Ⱥ�������Ⱥ���еı����仯��֪��S. metallicus��Ⱥ������ռ�����ı仯������Fe2+Ũ�ȵı仯���ƱȽ����ƣ���A. manzaensis��Ⱥ������ռ�����ı仯����Fe2+Ũ�ȵı仯�����෴���ڵ�9����Ʒ�У�Fe2+Ũ��������������������ߵ�(2.37 g/L)�������˵�15����Ʒ�У�Fe2+Ũ������0.62 g/L����ˣ����ƲFe2+Ũ�Ƚϸ�ʱ��S. metallicus�����ƾ��֣�����Fe2+�ϵ�ʱ��A. manzaensisռ���ƣ�����˵S. metallicus���к�ǿ����������������֮ǰ�ı���Ҳ����S. metallicus����������������ǿ��VILC?EZ��[26]�Ƚ���S. metallicus��M. sedula ��A. brierleyi����������ͭ���ʵ��������ΪS. metallicus������ǿ����������������M. sedula��֮����A. brierleyi���������о����������[26]�����Ľ�����������
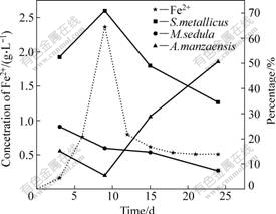
ͼ8 ����Һ��Fe2+Ũ�ȵı仯�����ֵı����仯����
Fig.8 Changing curves of Fe2+ concentration and percentages of each identified clone in solution
3 ����
1) ��ͭ��Ļ�Ͻ���ʵ��������������ȹž����нϸߵĽ���Ч�ʣ���pH 1.5��65 �������£�ҡƿ��Ͻ�����24��ʱͭ�������ܴﵽ90%���ϡ�
2) pHֵΪ 1.5ʱͭ������Ϊ90.8%����pH ֵΪ2.5ʱͭ������Ϊ56.9%����˵����һ����Χ�ڽϵ͵�pHֵ�������ڻ�ͭ��Ľ�����
3) ��pH 1.5�����£��¶ȴ�65 �����ߵ�75 ��ʱ����ͭ���ͭ������(87.7%)��δ���ӣ�˵���¶�����ʱͭ�����ʲ���һ����Ӧ���ߡ�
4) �ڻ�ͭ��Ľ���ǰ��S. metallicus��ռ�����Ƶľ��֣������˽�������A. manzaensis��Ⱥ������ռ������������������ȡ��S. metallicus��Ϊ�����֡�
REFERENCES
[1] Vilc��ez J, Yamada R, Inoue C. Effect of pH reduction and ferric ion addition on the leaching of chalcopyrite at thermophilic temperatures[J]. Hydrometallurgy, 2009, 96(1/2): 62?71.
[2] �� ƽ, �����, ������, �����. ��Ʒλԭ����ͭ�����������ǿ������[J]. ��ɫ����, 2004, 56(1): 34?36.
zou ping, ZHANG Shu-guang, ZHOU Xing-long, YANG Jia-ming. Intensified bioleaching of low grade chalcopyrite with thermoacidophile[J]. Nonferrous Metals, 2004, 56(1): 34?36.
[3] Norris P R, Owen J P. Mineral sulphide oxidation by enrichment cultures of novel thermoacidophilic bacteria[J]. FEMS Microbiology Reviews, 1993, 11(1/3): 51?56.
[4] ���ٲ�, ������, �½���. ��ͭ����������жۻ������о���չ[J]. ϡ�н���, 2006, 30(3): 395?400.
Shu Rong-bo, RUAN Ren-man, WEN Jian-kan. Review on passivation of chalcopyrite during bioleaching process[J]. Chinese Journal of Rare Metals, 2006, 30(3): 395?400.
[5] �ܺ鲨, ��ΰ��, ��Ө, �����, ������. �����������������о���Ӧ�ý�չ[J]. ������ɽ, 2006, 364(10): 5?8.
zhou Hong-bo, ZENG Wei-min, LI Ying, QIU Guan-zhou, LIU Jian-she. Advances in research and application of bioleaching process for sulfide ores using thermophilic microorganism[J]. Metal Mine, 2006, 364(10): 5?8.
[6] ������, ������, �� ��, ��䯵t, ������. ����������˾�������Ʒλ��ͭ��[J]. ��ұ����, 2007, 27(4): 25?30.
zhang yan-sheng, QIN Wen-qing, WANG Jun, REN Liu-yi, HE Ming-fei. Bioleaching of copper sulfide ore by Acidithiobacillus ferrooxidans and Acidithiobacillus thiooxidans [J]. Mining and Metallurgical Engineering, 2007, 27(4): 25?30.
[7] Rodriguez Y, Ballester A, Bl��zquezm L, Gonz��lez F, Mu?oz J A. New information on the chalcopyrite bioleaching mechanism at low and high temperature[J]. Hydrometallurgy, 2003, 71(1/2): 47?56.
[8] Konishi Y, Asai S, Tokushige M, Suzuki T. Kinetics of the bioleaching of chalcopyrite concentrate by acidophilic thermophile Acidianus brierleyi[J]. Biotechnology Progress, 1999, 15(4): 681?688.
[9] Stott M B, Watling H R, Franzmann P D, Sutton D C, Parker A J. Biohydrometallurgy: fundamentals, technology and sustainable development, Part A[C]//International Biohydrometallurgy Symposium. Amsterdam: Elsevier Science, 2001.
[10] Brierley J A. Biohydrometallurgy��This microbiologist��s perspective[C]//Biohydrometallurgy: From the Single Cell to the Environment (IBS 2007). Frankfurt: Trans Tech Publications Ltd, 2007.
[11] Rawlings D E, Johnson D B. The microbiology of biomining: development and optimization of mineral-oxidizing microbial consortia[J]. Microbiology, 2007, 153(2): 315?324.
[12] SILVERMAN M P, LUNDGREN D C. Study on the chemoautotrophic iron bacterium Ferrobacillus ferrooxidans: I. An improved medium and harvesting procedure for securing high cell yield[J]. Journal of Bacteriology, 1959, 77: 642?647.
[13] Hershberger K L, Barns S M, Reysenbach A L, Dawson S C, Pace N R. Wide diversity of Crenarchaeota[J]. Nature, 1996, 384(6608): 420?420.
[14] DeLONG E F. Archaea in coastal marine environments[J]. Proceedings of the National Academy of Sciences of the United States of America, 1992, 89(12): 5685?5689.
[15] Yang Yu, Wan Min-xi, Shi Wu-yang, Peng Hong, Qiu Guan-zhou, Zhou Ji-zhong, Liu Xue-duan. Bacterial diversity and community structure in acid mine drainage from Dabaoshan mine, China[J]. Aquatic Microbial Ecology, 2007, 47(2): 141?151.
[16] C��rdoba E M, Mu?oz J A, Bl��zquez M L, Gonz��lez F, Ballester A. Passivation of chalcopyrite during its chemical leaching with ferric ion at 68 ��C[J]. Minerals Engineering, 2009, 22(3): 229?235.
[17] Hackl R P, Dreisinger D B, Peters E. Passivation of chalcopyrite during oxidative leaching in sulfate media[J]. Hydrometallurgy, 1995, 39(1):25-48.
[18] Stott M B, Waliting H R, Franzmann P D, Sutton D. The role of ironhydroxy precipitates in the passivation of chalcopyrite during bioleaching[J]. Minerals Engineering, 2000, 13(10): 1117?1127.
[19] Mousavi S M, Jafari A, Yaghmaei S, Vossoughi M. Bioleaching of low-grade sphalerite using a column reactor[J]. Hydrometallurgy, 2006, 82(1/2): 75?82.
[20] Rawlings D E, Dew D, Plessis C D. Biomineralization of metal-containing ores and concentrates[J]. Trends in Biotechnology, 2003, 21(1): 38?44.
[21] Hita R, WANG Hong-mei, Bigham J M, Torrent J, Tuovinen O H. Bioleaching of a pyritic sludge from the Aznalc��llar (Spain) mine spillage at ambient and elevated temperatures[J]. Hydrometallurgy, 2008, 93(1/2): 76?79.
[22] ������, ̷����, ���˽�, ������, ������. ������������Ӱ������̽��[J]. ʪ��ұ��, 2004, 23(3): 113?121.
HU Kai-guang, TAN Kai-xuan, YANG Shi-jiao, LIU Guo-fu, LIANG Jian-long. Bacteria leaching mechanism and effect factors[J]. Hydrometallurgy of China, 2004, 23(3): 113?121.
[23] Fowler T A, Crundwell F K. Leaching of zinc sulfide by Thiobacillus ferrooxidans: experiments with a controlled redox potential indicate no direct bacterial mechanism[J]. Applied and Environmental Microbiology, 1998, 64(10): 3570?3575.
[24] Schippers A. Microorganisms involved in bioleaching and nucleic acid-based molecular methods for their identification and quantification[C]//Microbial Processing of Metal Sulfides, Netherlands: Springer, 2007: 3?33.
[25] Yoshida N, Nakasato M, Ohmura N, Ando A, Saiki H, Ishll M, Igarashi Y. Acidianus manzaensis sp. nov., a novel thermoacidophilic archaeon growing autotrophically by the oxidation of H2 with the reduction of Fe3+[J]. Current Microbiology, 2006, 53(5): 406?411.
[26] Vilc��ez J, Suto K, Inoue C. Bioleaching of chalcopyrite with thermophiles: Temperature�CpH�CORP dependence[J]. International Journal of Mineral Processing, 2008, 88(1/2): 37?44.
������Ŀ�������ص�����о���չ�ƻ�������Ŀ(2010CB630901)
�ո����ڣ�2008-11-14�������ڣ�2009-12-04
ͨ�����ߣ���ѧ�ᣬ�����ڣ���ʿ���绰��0731-88879815��E-mail��xueling0714@yahoo.com.cn
[2] �� ƽ, �����, ������, �����. ��Ʒλԭ����ͭ�����������ǿ������[J]. ��ɫ����, 2004, 56(1): 34?36.
[4] ���ٲ�, ������, �½���. ��ͭ����������жۻ������о���չ[J]. ϡ�н���, 2006, 30(3): 395?400.
[5] �ܺ鲨, ��ΰ��, ��Ө, �����, ������. �����������������о���Ӧ�ý�չ[J]. ������ɽ, 2006, 364(10): 5?8.
[6] ������, ������, �� ��, ��䯵t, ������. ����������˾�������Ʒλ��ͭ��[J]. ��ұ����, 2007, 27(4): 25?30.
[22] ������, ̷����, ���˽�, ������, ������. ������������Ӱ������̽��[J]. ʪ��ұ��, 2004, 23(3): 113?121.
