
���±�ţ�1004-0609(2008)02-0342-07
ϲ��������˾�YN12����ļ��������ӿ�����
�����ϣ������֣��� �����ųɹ���ѧ�ᣬ�����
(���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ����ɳ410083)
ժ Ҫ��
������������Ȫˮ���з����һ���ж�������������YN12��������̬�������������������Լ�16S rDNA���з������֤�����þ��������ϲ��������˾�(Acidithiobacillus caldus)���ؽ�������ʵ�������YN12�����3CdSO4?8H2O���г�ǿ���ԣ�����߳�ʼCd2+����Ũ�ȴ�4.8 g/L���ڴ˻����ϣ���������3CdSO4?8H2OŨ�ȣ�������Cd2+����Ũ�ȿɴ�31.5 g /L(�൱��3CdSO4?8H2O 210 g/L)���ڸ�����Cd2+����Ũ���£���������3������Ӧ��������YN12����������ٶȺ����������Ծ��ܵõ��ϺõĻָ���
�ؼ��ʣ�
ϲ��������˾���YN12�������ӿ������������Ũ����
��ͼ����ţ�Q 93���� ���ױ�ʶ�룺A
Identification and cadmium (��) resistance of strain YN12, Acidithiobacillus caldus
DING Jian-nan, ZHU Ruo-lin, KANG Jian, ZHANG Cheng-gui, WU Xue-ling, QIU Guan-zhou
(School of Resources Processing and Bioengineering, Central South University, Changsha 410083, China)
Abstract: A moderately thermophilic sulfur-oxidizing strain, YN12, was isolated from an acidic hot spring sample collected in Yunnan Province, southeastern China. Morphological, physiological and 16S rDNA sequence data suggest that YN12 belongs to the species, Acidithiobacillus caldus. YN12 exhibits a very high degree of resistance to cadmium (��), with an original maximum tolerated concentration (MTC) of 4.8 g/L and an ultimate MTC of 31.5 g/L after adapted by a gradually increased concentration of 3CdSO4?8H2O. Continuously cultured in the medium containing 210 g 3CdSO4?8H2O/L (equal to 31.5 g/L of Cd2+) for three generations, the strain is capable of restoring its growth and sulfur oxidation activity to an approximately normal level.
Key words: Acidithiobacillus caldus; strain YN12; cadmium resistance; maximum tolerated concentration
ϲ��������˾�(Acidithiobacillus caldus)��������˾���(Acidithiobacillus)��Ωһ���ʶ����Ⱦ�����1994����HALLBERG��LINDSTROM���롢��������[1]���������Ƕ���������õIJ�����ʶ���þ���������������ر����ڽ�����������������Ӧ���е�Ӧ��Խ��Խ�㷺[2-4]��ϲ��������˾���Ӧ�ã�����ʼ����Ժ�����������(��ɰ)������Ԥ����������þ��������ӵĿ��Դ�һ��ʼ�ͱ��ܹ�ע��Ŀǰ�������й�ϲ��������˾��Ŀ����о���Ҫ������ ��[5-7]���������ж��������ӿ��Ե��о��������١�������Ļ�ѧ��ɽ�Ϊ���ӣ����Ž���Ľ��У����������⣬�������������ж��������������������������������ͽ������[8-9]����ˣ��о�ϲ��������˾�����Щ���ӵĿ��ԣ�������������Ч�ʾ���ʮ����Ҫ�����塣�������߷��봿����21��ϲ��������˾��¾��꣬���а���һ��߿���(Cd2+)����YN12�����ı����˸þ���ķ��롢�������俹Cd2+���ܵ��о������
1 ����ķ��������
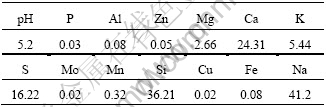
���������ɼ���������Ȫˮ��һʽ2�ݣ�����һ�ݱ�����45 �棬���ھ��ָ�������һ�ݱ�����4 �棬����Ԫ�ط�������Ʒ�Ӳɼ���ʹ�ã�����ʱ�䲻����4 d������ԭ�����շֹ��ȷ�(Atomic absorption spectrometry��AAS)����ˮ����ѧ�ɷ֡���������Ȫˮ������Ҫ���Լ���1��
��1 ������Ȫ��Ʒ80 ��ʱ����Ҫ����
Table 1 Main properties of acidic hot spring sample at 80 ��
��3 mL 45 ���±����ˮ���ӵ�װ��47 mL����������������ƿ�У����������pHֵ��2.5��Ȼ��������������(S0)����45 �桢ת��180 r/min��������5~7 d��ÿ����pHֵ�仯����pHֵ����1.0����ʱ����ͬ������ת����������pHֵ�ٴν���1.0����ʱ��������һ��ת����������˸���3�Ρ���3�θ������������о�����롣����������[10]�������»����γɷ�(g/L)��(NH4)2SO4 (3.0)��Na2SO4?10H2O (3.2)��KCl (0.1)��K2HPO4 (0.05)��MgSO4?7H2O (0.5)��Ca(NO3)2 (0.01)������0.02%(w/v)��ĸ��ȡ��(YE)��������������������Ԫ�أ���Ԫ�ص��䷽����(mg/L)��FeCl3?6H2O (11.0)��CuSO4?5H2O (0.5)��HBO3 (2.0)��MnSO4?H2O (2.0)��Na2MoO4?2H2O (0.8)��CoCl2? 6H2O (0.6)��ZnSO4?7H2O (0.9)��Na2SeO4 (0.1)����������������YE���ó����ѹ������������Ԫ�ع��˳�����S0���ü�Ъ������������3�Ρ�
����3�θ���������Ϳ�����䷽�븻����������ͬ���������ʲ�ͬ��pHֵΪ2.5��0.7%(w/v)��֬ƽ���ϡ��ù��������������������ɸ�����������S0��Ϊ1.5 g K2O6S4/L(���˳���)����Ϳ���õ�ƽ������45 ��������7 d����ȡ���������������������������ͬ��pH 2.5��Һ���������У�������5~7 d��Ȼ������ǰ���������������룬��˷�������3�Σ����õ���������YN12���ꡣ�þ�����4 ���±�����Һ�帻���������У����ڻ��������˵���⣬����ʵ��������������Ϊ����������
1.2.1 16SrRNA�������
��YN12����������Һ���������У������������ﵽ�ȶ���ʱ������������ֽ����2�Σ���ȥ��ۣ�ȡ100 mL��Һ�������ռ���ϸ�����Ⱥ���pH 1.8ֵΪ���ҵ�ϡ������Һ��pHֵΪ8.0��TE����Һ�ֱ�ϴ��2�Σ�Ȼ��������200 ��L TE����Һ�С�ʹ��EZ-10 Spin Column Genomic DNA Minipreps Kit (Bio Basic Inc.)�Լ��У���˵����ȡ������DNA����63f(5��- CAGGCCTAACACATGCAAGTC-3��)/1387r (5��-GGG- CGGWGTGTACAAGGC-3��)����ԣ�����YN12�����16S rDNA���С�PCR��ϵ25 ��L��������2.5 ��L 10��PCR ����Һ��2.5 ��L (25 mmol) MgCl2��1 ��L (10 mmol) dNTP mix (Fermentas)��1 ��L 63f�������ͬ�����1387r���1 ��L (5 units) Taqø (Fermentas)��1 ��Lģ��DNA �Լ�15 ��L ���˫��ˮ��������ϵ��ģ��DNA����ͬ��������˫��ˮ�����⣬������ǰ����ϵ��ȫ��ͬ��PCR�������£�94 ���ʼ����5 min��33��ѭ����ÿ��ѭ������94 �����45 s��55 ���˻�45 s��72 ��������90 s��ѭ�����������72 ��������10 min��PCR����ʹ��E.Z.N.A? Gel Extraction Kit (Omega Bio-Tek, Inc.)�����Լ��д������������PCR������pBS-T PCR Products Clone Kit (Tiangen Biotech Co., Ltd., Beijing)��¡�Լ��п�¡ת�������巽�������Ľ�����[11]���о��ɹ��������Կ�¡��Һ�ͱ�������Զ־���\���������ι�˾������16S rDNA����
ʹ��Clustal X (1.8) �� MEGA3��������16S rDNAϵͳ���������Ա���YNTC-1������������������Ե��ϵ��
1.2.2 ������̬����
�����崦���ȶ��ڵ�YN12��Һ����������ֽ���ˣ��ռ���ϸ��������pHֵΪ2.5���ҵ�ϡ����ϴ��2�Σ�ȡ����ϸ����Ƭ����������Ⱦɫ���ڹ�ѧ�����¹۲죬ʣ��ϸ���ͺ���ʦ����ѧ������ѧѧԺ�羵�ҽ���ɨ��羵������
1.2.3 ����������������
��YN12���������Һ���������У��ֱ�����30��35��40��45��50��55��60 ����¶�����������ʼpHֵΪ2.5������100 h����ֱ�Ӽ���������Ϲ��ܶȣ�����������״���������¶ȷ�Χȷ��������С�¶ȼ����������ͬ���ķ�ʽ����ʵ�飬ֱ����ȷ����1 �档
������ͬ���ķ�ʽ���֣���ϡ����ֱ������������ʼpHֵΪ1.0��1.5��2.0��2.5��3.0��3.5��4.0��4.5����YN12����������¶�������100 h��������ͬ���ķ����۲�������״����
��YN12��Һ���ģ��ռ���ϸ������pHֵΪ2.0���ҵ�ϡ����ϴ��2�Σ�����ͬ�����������ڷֱ���S0(5 g/L)��Na2S2O3(10 g/L�����˳���)��K2O6S4(1.5 g/L�����˳���)��FeSO4?7H2O (5 g/L�����˳���) ��������(2 g/L)��YE(0.2 g/L)��������(1 g/L)��S0(5 g/L)+������(1 g/L)��S0(5 g/L)+YE(0.2 g/L)��S0(5 g/L)+������(1 g/L)Ϊ��Դ���ʵ�Һ���������У���YN12�����������ʼpHֵ�������¶Ⱥ�ת��180 r/min�£�������100 h����ֱ�Ӽ���������Ϲ��ܶȣ����������������������Դ�������������
2 ������ӿ���
ʵ��ǰ�Ƚ�YN12���������ǰ��S0�������У��������������(��2)�»����pHֵ����1.0ʱ����ҡ����ȡ���������¾���24 h����һϵ��װ��47 L ��ͬ������������ƿ�зֱ����3 L��ľ�Һ����Щ����ƿ�к���ͬŨ�ȵķ�����3CdSO4?8H2O��ʵ������ͬpHֵ(2.5)����ͬ��ҡ�ٶ�(180 r/min)�Լ�����������¶��½��С�������10��Ũ�ȣ�����һ�飬���а���0 g 3CdSO4?8H2O/L�Ķ��ա��ֱ�ȡ��ʵ��ƿ��Һ��UV 2300�ֹ��ȼ�(Techcomp)�ϣ��ڲ���440 nm�����ⶨ��ʼ���ܶ�(Optical Density, OD)ֵ����ø�ʵ��ƿ����ʼODֵΪ0.020~.025����ȡ3ƿ��Һ����ϸ����������ø����ƽ����ʼ��ϸ����Ϊ2.1��106 /mL��ÿ��ʵ���ظ�һ�Ρ�ʵ������ж��ڲⶨ������վ�Һ��pHֵ����pHֵ����1.0ʱ��ʵ��ֹͣ�������ⶨ��������ʵ��ƿ��pHֵ������ʵ���¶��¾���3 h��ȡ��ƿ��Һ����Һ������������CS2���ܽ���������ھ�Һ�е�Ԫ���������ⶨ����ODֵ�����Ӹ�ʵ��ƿ��ȡ������Һ�����ʵ�ϡ�ͺ���Ѫ�������ֱ�Ӽ���������Olympus CX31��ѧ�����²ⶨ���ջ�ϸ������������ODֵ����ʼODֵƽ����0.005�����ջ�ϸ��ƽ���ﵽ2.5��106/ mLʱ��ʵ��ƿ�е�Cd2+Ũ�Ⱦ��Ǹþ����Cd2+����߳�ʼ����Ũ��(Original maximum tolerated concentration, OMTC)��
δ��Cd2+ѱ����YN12�����Ѷ�Cd2+���нϸߵij�ʼ���ԣ�Ϊ�о��俹Cd2+�����Ƿ���ͨ��Cd2+ѱ���õ���һ������ߣ����ǴӾ������߳�ʼCd2+����Ũ��(4.8 g/L)����YN12����������Cd2+Ũ�ȵĿ���ѱ����ѱ�������ж��ڲⶨ��ҺODֵ���������pH���۲���������Cd2+Ũ�ȶԸþ����������������Ե�Ӱ�졣3CdSO4?8H2OŨ�ȵľ�����������Ϊ��32 g/L��64 g/L��96 g/L��120 g/L��144 g/L��173 g/L��210 g/L��ÿ��Ũ�Ⱦ��費��3CdSO4? 8H2O �Ķ��գ��������ֱ��ظ�3�Σ���������������߳�ʼ����Ũ��ʵ����ͬ��ÿ�ν���ǰ���ռ�ǰһ��Cd2+Ũ���������ľ�ϸ����������ˮϴ��2�Σ�Ȼ���ٽ���ʵ��ƿ�У�����ƿ���֣���ǰ�ζ��������������ľ�ϸ��������������ʵ��ƿ��������ȫ��ͬ������ƿ��ʵ��ƿ�Ľ�����Ҳ��ȫ��ͬ��ODֵ���������pH�ⶨ��������߳�ʼ����Ũ��ʵ����ͬ����ǰ�����Σ����24 h��ODֵ�ͻ�����������仯��Ϊʵ����ֹ�����ݣ���ʱ��ODֵ�ͻ����������Cd2+Ũ���µ����վ�Ũ�ȡ�
ʵ������ν��С���һ���ǰ���������������Cd2+Ũ�ȵ�����ѱ���Σ�Ŀ����ȷ������Cd2+Ũ����YN12��Һ�ܴﵽ������Ũ�ȣ��Լ��ﵽ�þ�Ũ������Ҫ��ʱ�䣻�ڶ�������ѱ���������������о��Σ��ý��ǽ���������Cd2+Ũ��ѱ���ĵ�һ��YN12���꣬�����ڴ�Cd2+Ũ����ת��2�Σ��õ�ѱ���ڶ����͵��������꣬�۲�YN12����������ٶȡ����վ�Ũ�Ⱥ�pH�Ƿ������ת�Ӵ��������Ӷ������仯��
3 ���������
�Բɼ���������Ȫˮ��������3�θ������ٶԵ�3�θ������������3����֬ƽ����룬�õ���������YN12���ꡣ
YN12���������Ⱦɫ��ӦΪ���ԣ���ѧ������ɨ��羵��ʾ�þ�Ϊ���͵Ķ̸�״����СΪ0.5~0.6 ��m�� 1.3~1.4 ��m����ͬ�����ȶ��ڵ�ͬ������20�����Ƚϣ�YN12ϸ���϶�(���������СԼ0.4~0.6 ��m��1.4~1.8 ��m)�����ͼ1�ͱ�2��
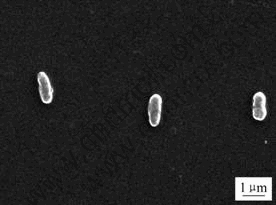
ͼ1 YN12����ɨ��羵��Ƭ
Fig.1 SEM image of strain YN22 (��10 000)
��2 YN12�������Ҫ����
Table 2 Main properties of strain YN12
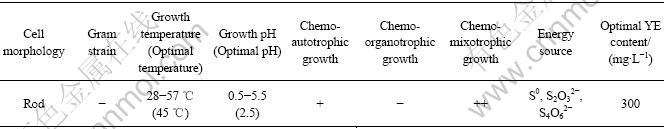
��2�г���YN12�������Ҫ��̬�����������������ԡ��þ���������¶�Ϊ28~57 ��(�����¶�Ϊ45 ��)������pHֵΪ0.5~5.5(����pHֵΪ2.5)��������S0��S2O32-��S4O62- ���л����������������ܽ���ר����������������S0Ϊ��Դ���ʵ�����¿��Խ��л��Ӫ������������������£������ˡ�YE�������ǿ��Դٽ���������������YEŨ��300 mg/L��ͼ2��ʾΪ��ͬ��Դ������YN12���������������þ��겻������Fe2+��S0Ϊ��Դʱ������ã���ĸ��ȡ����������ٽ���������
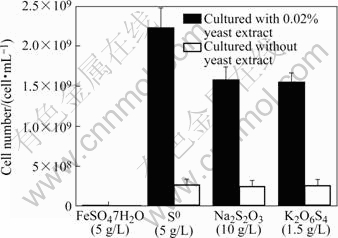
ͼ2 ��ͬ��Դ���ʶ�YN12����������Ӱ��
Fig.2 Effect of energy source on growth of YN12
����NCBI�е�BLAST(blastn)�����YN12�����16S rDNA����Ƭ����GenBank����¼��16S rDNA���н���ͬԴ�ԱȽϣ�����YN12�����16S rDNA������GenBank�����е�ϲ��������˾�����(Acidithiobacill caldus) 16SrDNA���е������Զ���99%���ϡ�ͼ3��ʾΪʹ��3.1�汾MEGA����[12]�����Ļ���16S rDNA���е�ϵͳ��������������YN12������ϲ��������˾������� DSM 8584������ͬ�־������ͬһ�飬��������������˾�(A. ferrooxidans)�����������������˾�(A. thiooxidans)������ֱ�����������顣
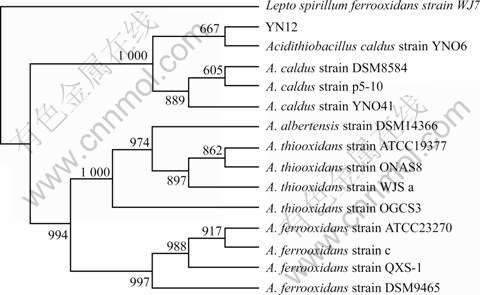
ͼ3 ����16SrRNA�������е�ϵͳ������
Fig.3 Phylogenetic tree based on 16S rRNA gene sequences of strain YN12 and related species
YN12�������Ҫ��̬�����������������ԣ��Լ�16S rDNA���з�����������ѱ�����ϲ��������˾�������ͬ[1, 13-15]���о����֤��YN12���������ϲ��������˾���
3.2.1 ��ʼCd2+����Ũ��
YN12�����3CdSO4?8H2O����߳�ʼ����Ũ��Ϊ32 g/L����ʵ��Cd2+Ũ�ȴﵽ4.8 g/L�������������4~32��(�������������δ�г�)�����ֳ���ǿ�Ŀ�Cd2+���ܡ���ͼ4���Կ���0.5~2.0 g/L��3CdSO4?8H2OŨ�ȶ�YN12��������û������Ӱ�죬����3CdSO4?8H2OŨ�����ӵ�4 g/Lʱ��YN12��Һ�����չ��ܶ�ֵ��ͬ�ڵĶ��վ�Һ�����25.1%����������ֵ�������о�����3CdSO4?8H2OŨ�����ӵ�8.0 g/Lʱ����ʼ��YN12���������������ã�����3CdSO4?8H2OŨ�ȵ����ӣ���YN12�������������ý�һ����ǿ����16 g/Lʱ��ODֵ�ȶ����½���22.2%���ڴ˻����Ͻ�3CdSO4?8H2OŨ�������100%���ﵽ32.0 g/Lʱ����ϸ���Ѳ���������ֳ����ʱ��ODֵֻ����ʼODֵ��0.005�������Ҳֻ��2.5��106 /mL�����Ը�����ʼֵ(2.1��106 /mL)���������Ũ������YN12��3CdSO4?8H2O����߳�ʼ����Ũ�ȡ�
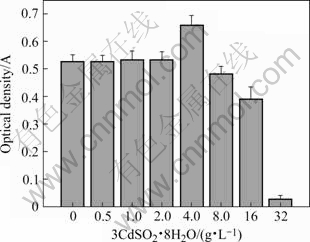
ͼ4 3CdSO4?8H2O��YN12����������Ӱ��
Fig.4 Effect of 3CdSO4?8H2O on growth of strain YN12
3.2.2 Cd2+����ѱ��
ͼ5��ʾΪ��ͬCd2+Ũ���£�YN12��������Һ�ܴﵽ�����ODֵ��������ʱ�䡣����Cd2+Ũ�ȵ���������ODֵ�����½����Ӷ�����Ʒ(0 g/L 3CdSO4?8H2O)��0.536��������Cd2+Ũ��(210 g/L 3CdSO4?8H2O)�µ�0.313����64 g/L��96 g/L 3CdSO4?8H2O֮���ODֵ�½��������ͬʱ������Cd2+Ũ����ߣ��ﵽ��ӦCd2+Ũ�������ODֵ������ʱ��Ҳ�����ӣ�3CdSO4?8H2O��Ũ��Ϊ0 g/Lʱ���ﵽ0.536�����ODֵֻ��100 h������3CdSO4?8H2OΪ210 g/L������Ũ��ʱ����Ҫ288 h���ܴﵽ���ODֵ0.313��
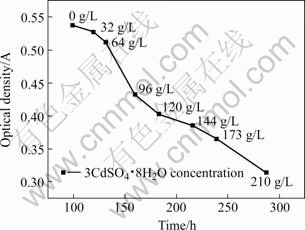
ͼ5 ��ͬ3CdSO4?8H2OŨ�ȶ�YN12����������Ӱ��
Fig.5 Effect of various 3CdSO4?8H2O concentrations on growth of strain YN12
��������Ũ��Cd2+(210 g/L 3CdSO4?8H2O)ѱ����YN12����������ͬCd2+Ũ����ת�������������ٶȳ�����ߵ�����(ͼ6)���������ODֵ��ѱ��һ��(210 g/L 3CdSO4?8H2O-��)��0.313��ߵ�ѱ������(210 g/L 3CdSO4?8H2O-��)��0.4����������ߵ�ѱ������(210 g/L 3CdSO4?8H2O-��)��0.46�����Ҵﵽ��߾�Ũ�ȵ�ʱ��Ҳ��288 h(ѱ��һ��)���̵�240 h(ѱ������)��192 h(ѱ������)����pHֵ�ı仯�������������ѱ�����������ӣ�Խ��Խ�ӽ���Cd2+������Ʒ(ͼ7)��˵���������������������ָ���YN12����������pHֵΪ2.5����Ac base-S0������������ʱ������S0��������SO42+�����ۻ���pHֵ�����½������͵�pHֵ������ϸ����������ֳ��������������ϸ�����ܡ����ͼ6��ͼ7����������ؿ��������о�Һ������Ũ�Ⱦ�������pHֵΪ1.0����������pHֵ�����½�����Ũ�Ȳ��Ͻ��͡���ʵ�����ڣ���Ϊ���վ�Һ��pHֵ�½���������½��ٶ���죬���Ծ�Ũ���½�Ҳ��Ѹ�٣��෴��ѱ��һ����Һ��pHֵ�½���Ϊ���ͣ��������ʵ�����ʱ��Ũ����δ�������ԵĽ��͡�

ͼ6 ����Cd2+Ũ��������������YN12������������
Fig.6 Growth curves of strain YN12 continuously grown at final Cd2+ concentration
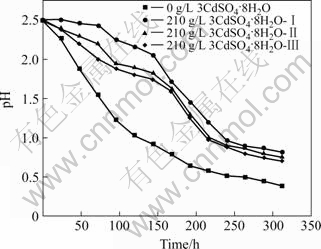
ͼ7 ����Cd2+Ũ��������������YN12��Һ��pHֵ���ݻ�
Fig.7 pH evolution in cultures of strain YN12 continuously grown at final Cd2+ concentration
YN12�����Cd2+�Ŀ���ʮ��ͻ����δ��ѱ���ľ����Cd2+����߳�ʼ����Ũ�ȴﵽ4.8 g/L(32.0 g/L 3CdSO4?8H2O)����������Cd2+Ũ�ȵ�ѱ��ʵ���һ��������YN12���������Cd2+����Ũ�ȿ��Ըߴﺱ����31.5 g/L(�൱��210 g/L 3CdSO4?8H2O)���俹Cd2+����ԶԶǿ�����������ǽ�������[16-18]��YN12��Ŀǰ�������Ŀ�Cd2+������ǿ�ľ��ꡣ
������Cd2+����Ũ��(31.5 g/L)�£���ѱ�����YN12�����������ת�������������ʾ�þ��ڸ�Cd2+Ũ���£�����������������Ӧ���������������ٶȺ����������Ծ��õ��˽ϺõĻָ���˵���þ����н�һ�����Cd2+���Ե�DZ����
��Ȥ���ǣ�YN12��Դ�ڲ������ӵ�������Ȫ(��1)������Cd2+ȴ������˸ߵĿ��ԣ����Cd2+�Ŀ��Ի���ֵ�������о������⣬�þ��ڸ�����ӹ������Ӧ��ǰ��Ҳֵ������̽�֡��������߽��ڴ˻����ϣ���YN12���꿹Cd2+�����������ͷ�������ѧ���ƣ��Լ��þ��ڸ�����ӹ����濪չ��һ���Ĺ������Գ��������俹Cd2+������Ӧ��DZ����
4 ����
1) �Բ������ϵ�������Ȫˮ������3�ζ�����Ȼ��Ե�3�θ����������֬ƽ����룬�õ��ʶ������������������䣬�õ������پ�����ƽ����룬���õ���������YN12���꣬�þ��꾭��̬ѧ�۲졢���������о���16S rRNA�������з�����֤��Ϊϲ��������˾���
2) ʹ�÷�����3CdSO4?8H2O��δ��Cd2+ѱ����YN12������еij�ʼCd2+����Ũ��ʵ���������Cd2+��߳�ʼ����Ũ�ȴ�4.8 g Cd2+/L(�൱��32 g/L 3CdSO4?8H2O)����������Cd2+Ũ�ȵ�ѱ��ʵ���һ��������YN12�����Cd2+�Ŀ��Կ���ͨ��ѱ��������ǿ�����Cd2+����������Ũ�ȴﵽ������31.5 g/L(�൱��210 g/L 3CdSO4?8H2O)��
3) ��31.5 g/L������Cd2+����Ũ���£���YN12��������������������ָþ��ڸ�Cd2+Ũ���£�����������������Ӧ���������������ٶȺ����������Ծ��ܵõ��ϺõĻָ���˵���þ����н�һ�����Cd2+���Ե�DZ����
4) YN12������Դ�ڲ������ӵ�������Ȫ������Cd2+ȴ������˸ߵĿ��ԣ���Cd2+���Ի���ֵ�������о���ͬʱ���þ��ڸ�����ӹ�����Ҳ���нϺõ�Ӧ��ǰ����ֵ������̽�֡�
REFERENCES
[1] HALLBERG K B, LINDSTROM E B. Characterization of Thiobacillus-Caldus sp-nov, amoderately thermophilic acidophile[J]. Microbiology, 1994, 140: 3451-3456.
[2] DOPSON M, LINDSTORM E B. Potential role of Thiobacillus caldus in arsenopyrite bioleaching[J]. Appl Environ Microbiology, 1999, 65(1): 36-401.
[3] SEMENZA M, VIERA M, CURUTCHET G, DONATI E. The role of Acidithiobacillus caldus in the bioleaching of metal sulfides[J]. Latin American Applied Research, 2002, 32: 303-306.
[4] OKIBE N, JOHNSON D B. Biooxidation of pyrite by defined mixed cultures of moderately thermophilic acidophiles in pH-controlled bioreactors: Significance of microbial interactions[J]. Biotechnology and Bioengineering, 2004, 87(5): 574-583.
[5] �� ��, ����÷, ղ ��, �ֽ�Ⱥ, ������. һ���Ч����ϲ����˾����̾��Ĺ���[J]. ����ѧ��, 2005, 45(5): 675-679.
ZHAO Qing,��LIU Xiang-mei,��ZHAN Yang,��LIN Jian-qun, YAN Wang-ming. Construction of an engineered Acidithiobacillus caldus with high efficiency arsenic resistance[J]. Acta Microbiologica Sinica, 2005, 45(5): 675-679.
[6] TUFFIN I M, HECTOR S B, DEANE S M, RAWLINGS D E. Resistance determinants of a highly arsenic-resistant strain of Leptospirillum ferriphilum isolated from a commercial biooxidation tank[J]. Applied and Environmental Microbiology, 2006, 72(3): 2247-2253.
[7] KOTZE A A, TUFFIN I M, S M DEANE, RAWLINGS D E. Cloning and characterization of the chromosomal arsenic resistance genes from Acidithiobacillus caldus and enhanced arsenic resistance on conjugal transfer of ars genes located on transposon TnAtcArs[J]. Microbiology, 2006, 152(12): 3551-3560.
[8] SILVER S, SCHOTTEL J, WEISS A. Bacterial resistance to toxic metal ions��A review[J]. Gene, 1996, 179(1): 9-19.
[9] RAWLINGS D E. Heavy metal mining using microbes[J]. Annu Rev Microbiol, 2002, 56: 65-91.
[10] DOPSON M, LINDSTROM E B. Potential role of Thiobacillus caldus in arsenopyrite bioleaching[J]. Applied and Environmental Microbiology, 1999, 65(1): 36-40.
[11] XIA Jin-lan, PENG An-an, HE Huan. A new strain Acidithiobacillus albertensis BY-05 for bioleaching of metal sulfides ores[J]. Trans Nonferrous Met Soc China, 2007, 17(1): 168-175.
[12] KUMAR S, TAMURA, K, NEI M. Integrated software for molecular evolutionary genetics analysis and sequence alignment[J]. Briefings in Bioinformatics, 2004, 5(2): 150-163.
[13] OKIBE N, GERICKE M, HALLBERG K B, JOHNSON D B. Enumeration and characterization of acidophilic microorganisms isolated from a pilot plant stirred-tank bioleaching operation[J]. Applied and Environmental Microbiology, 2003, 69(4): 1936-1943.
[14] KAMIMURA K, OKAYAMA T, MURAKAMI K, SUGIO T. Isolation and characterization of a moderately thermophilic sulphur-oxidizing bacterium[J]. Microbios, 1999, 99(392): 7-18.
[15] �� ӧ, ��ž�, �ֽ�Ⱥ, �����, ������. һ���ж����������������˾��ķ����ϵͳ��������[J]. ����ѧ��, 2004, 44(3): 382-3851.
LIU Ying, QI Fang-jun, LIN Jian-qun,��TIAN Ke-li, YAN Wang-ming. Isolation and phylogenetic analysis of a moderately thermophilic acidophilic sulfur oxidizing bacterium[J]. Acta Microbiologica Sinica, 2004, 44(3): 382-3851.
[16] ��, ������, ������. һ�꿹�ؽ���ͭ��ϸ���ķ��롢��������16SrDNA�����з���[J]. ����ѧͨ��, 2005, 32(3): 68-72.
PAN Yuan-yuan,��CHEN Wen-li,��HUANG Qiao-yun. Isolation, identification and16SrDNA sequences analysis of a bacterial resistant to copper and cadmium[J]. Microbiology Aviso, 2005, 32(3): 68-72.
[17] ABOU-SHANAB R A I, VAN BERKUM P, ANGLE J S. Heavy metal resistance and genotypic analysis of metal resistance genes in gram-positive and gram-negative bacteria present in Ni-rich serpentine soil and in the rhizosphere of Alyssum murale[J]. Chemosphere, 2007, 68: 360-367.
[18] SHENG Xia-fang, XIA Juan-juan. Improvement of rape (Brassica napus) plant growth and cadmium uptake by cadmium-resistant bacteria[J]. Chemosphere, 2006, 64: 1036-1042.
������Ŀ�������ص�����о���չ�ƻ�������Ŀ(2004CB619201); ������Ȼ��ѧ�������о�Ⱥ���ѧ����������Ŀ(50621063)
�ո����ڣ�2007-05-08�������ڣ�2007-11-04
ͨѶ���ߣ������, ����, ��ʿ; �绰: 0731-8879212; E-mail: guanzhouqiu@yahoo.com.cn
