
�Ǿ���ûʳ����������������Ʊ�����Ե�ˮ��������Ӱ��
����1, 2������ӿ1, 2��³����1, 2����ΰ1, 2�������1, 2
(1. ���ȴ�ѧ ����ѧԺ��������ѧ�빤��ϵ������ ���ţ�361021��
2. ���ȴ�ѧ ��������Դ�����о��������� ���ţ�361021)
ժ Ҫ��
���Ǿ���(CTS)����ûʳ����(GA)��Ӱ�����أ������ó��Ǿ��Ƕ�ûʳ�����������������ǣ�ûʳ������Һ��ʼ����Ũ��Ϊ1 200 mg/L��20 mLûʳ������Һ�пǾ�������Ϊ0.02 g����ʱ��Ϊ60 min���¶�Ϊ25 �档�ڴ��������Ʊ���ûʳ������Ǿ��ǵ�������Ϊ623 mg/g������������о�������ͷ��ʡ�Ȼ��̽�ָ��������ˮ������(Microcystis flos-aquae)�͵���С����(Chlorella pyrenoidosa)������Ӱ�졣�о�������������������������ˮ����������������������ã������������Ũ��Ϊ30 mg/Lʱ������ʱ�佫�ﵽ15 d�������������Ũ��Ϊ60��90��130 mg/Lʱ������ʱ�佫����24 d�������ʸߴ�99%��������15 d�ڣ��������δ�Ե���С������ֳ����Ե��������ã������������Ũ��Ϊ60 mg/Lʱ�������ʽ�Ϊ5%����ˣ��Ǿ�����ûʳ�������������������Ч����ˮ������ˮ��������������Ե���С�����Ӱ�������
�ؼ��ʣ�
�������������ûʳ������ˮ������������С������
��ͼ����ţ�X524 ���ױ�־�룺A ���±�ţ�1672-7207(2014)07-2538-09
Effects of new algaecide chitosan-gallate on growth of Microcystis flos-aquae and Chlorella pyrenoidosa
LIU Yang1, 2, GUO Peiyong1, 2, LU Beibei1, 2, HUANG Wei1, 2, WAN Jinjin1, 2
(1. Department of Environmental Science and Engineering, College of Chemical Engineering,
Huaqiao University, Xiamen 361021, China;
2. Institute of Environmental and Resources Technology, Huaqiao University, Xiamen 361021, China)
Abstract: The factors of chitosan adsorbed gallic acid in orthogonal experiments were studied. The best adsorption condition was during shaking for 60 min at 25 �� with initial concentration of gallic acid of 1 200 mg/L and 0.02 g chitosan in 20 mL gallate acid. The adsorption ratio of chitosan to gallate algaecide of 623 mg/g was prepared under this condition and the short-term release rate of this algaecide as well as its effects on the growth of Microcystis flos-aquae and Chlorella pyrenoidosa were studied. The results show that the inhibitory effect on the growth of Microcystis flos-aquae can last up to 15 d when the algaecide mass concentration is 30 mg/L; when the algaecide mass concentration is 60, 90 and 130 mg/L, the inhibition time is even longer than 24 d with 99% inhibition rate. However, this algaecide does not show significant effects on Chlorella pyrenoidosa within 15 d. When the algaecide mass concentration is 60 mg/L, the inhibition is only 5%. Therefore, this algaecide can control the growth of Microcystis flos-aquae effectively, while it has weak influence on Chlorella pyrenoidosa.
Key words: algaecide; sustained-release; gallic acid; Microcystis flos-aquae; Chlorella pyrenoidosa
��������ˮ�帻Ӫ��������������ˮ�����������Ͼ���ˮ��ľ��ۺ���̬�����ܵ�����Ӱ��[1]�����ڶ�����ˮ�����о��У����û����������������������������オ���Ժ���̬��ȫ�Զ������ܵ����ӡ����������ǻ������õ����壬�����������ǻ��Խ�ǿ��һ�������[2]������ˮ���ᡢûʳ����Ͷ�����ձ�����ڶ���ֲ��Ļ��������У��ڽϵ�Ũ�Ⱦ��н�ǿ�Ļ������ã��������ѧ���Ѿ�������벢���о������Ƕ����������ֲ�����������[3-4]��Kamaya��[5-7]���о��������ˮ����Լ�������������Ե�����Ч����Wang��[8]֤��ûʳ����Ȼ������ʶ�ͭ����������������Ե��������á����ǣ���Щ�о�������ģ��ˮ�廷����ֱ��Ͷ�ŷ���������ʣ�������ɾֲ�Ũ��̫�߶������������������Ӱ�죬�����������Чʱ��Ҳ�϶̡���ˣ����þ��л����ܵ������������ˮ���������������ܳ�Ϊ����ˮ������Ч�ķ������Ǿ���(Chitosan)����Ȼ��Ψһ�ļ��Զ��ǣ�������һ����Դ�ḻ�������オ�����Ȼ�ۺ���[9]�������[10]���ü��ء��Ǿ��ǵ�������������ˮ��Һ��ͭ���Ӻ�Գೱ�����ɱ��Ϳ������ã�������ֱ��Ͷ������ͭ�����ľֲ�Ũ�ȸ߶��˺������ȱ�㡣����ɵ�[11]�ԿǾ���������ϳɻ����ԵĿǾ�����ͭ�����������Ч���Ƶ���С�����������������������������ӣ�����������ͬʱ���Ǿ�����Ϊ������������ˮ���ؽ���������к����ʵ�ȥ���Լ�����ˮ��Ҳ���˽϶���о�[12-14]�������������������������Ϊ���Ƽ����������û��������Ѿ�����һ���о������ǻ�������������Ũ�ȷֲ������ͳ���ʱ�䲻������⣬���ԿǾ����������������Ʊ����������δ����������ˣ����������ԿǾ��Ƕ�3�ֻ�������ˮ���ᡢ����Ӻ�ûʳ��������������о�Ϊ����㣬ɸѡ�����������Ⱥ������ʽϸߵ���ϣ��Ʊ����ø�Ч��������������������ˮ��������г��ڵ�����ʵ�飬���Ե���С������Ϊ�Աȣ��Ӷ�Ϊ��Ч����ȫ�Ŀ�������������ֳ����ֹ������ˮ�������ṩ�ɿ��Ŀ�ѧ���ݡ�
1 �����뷽��
1.1 ʵ�����
ˮ������(Microcystis flos-aquae)(FACHB-1028)Ϊˮ�����ֲ����ͷ��嶾�أ�����С����(Chlorella pyrenoidosa)(FACHB-9)Ϊ������Դ���֣���2����������й���ѧԺ�人ˮ�������о�����ˮ���ֿ⣬���ҷֱ�ʹ��ָ����������BG11��������Bristol����������������ʵ��ǰ�������ڵ����ֽ��ֵ�װ��400 mL������������ 1 L����ƿ������������ϸ���ܶȴﵽ����������ʱ��װ��100 mL��������Ŀ���ƿ�У��ȶ�1 d����к���ʵ�顣�����¶�Ϊ (25��1) �棬�ⰵ��Ϊ12 h:12 h��ÿ��ҡ��3�Σ����������λ�ã�����ʹ��ƿ�Ĺ�ͨ������һ���Լ�СżȻ��
�Ǿ��ǺͶ����(�ڱ�����)���Թ�ҩ���Ż�ѧ�Լ�����˾��ˮ���Ṻ�Թ㶫��ͷ����¤������ûʳ���Ṻ�Ի����ơ��Ǿ����������ȣ�80%������ӡ�ˮ�����ûʳ�����Ϊ��������
1.2 �Ǿ��Ƕ�3�ַ�������������������о�
��������ֹ��ȼ�ɨ��ˮ���ᡢ����Ӻ�ûʳ����3�ַ�������չ��ף��������շ壬Ȼ�����Ũ���������صı����ߡ�ȡ0.05 g�Ǿ��Ƿֱ����20 mL��ʼŨ��Ϊ100��200��300��400��500��600��800��1 000��1 200��1 500 mg/L��3�ַ�����Һ�У�Ȼ����25 ��ˮԡ������l h���˺�����Ӧ�����²ⶨ������ȼ�����Һ��δ�������ķ����Ũ�ȣ���������������ʺͿǾ��ǵ������Ⱥ������ʡ�����������£��Ǿ��Ƕ�ûʳ�������������ߣ�ûʳ����Ũ��Ϊ1 000 mg/Lʱ�������ʿ��Դﵽ85.7%����Ӧ������Ϊ491 mg/g���Ǿ��Ƕ�ˮ����Ͷ���ӵ���������ʷֱ�Ϊ43.3%��14.5%����Ӧ�����ȷֱ�Ϊ138 mg/g��46 mg/g������Ч����Խϲ����ѡȡûʳ������к���ʵ�顣
1.3 �Ǿ�����ûʳ������������Ʊ�
�ֱ��о�ûʳ������Һ��ʼŨ�ȡ��Ǿ��dz�ʼ����������ʱ����¶ȵȵ����������¿Ǿ��Ƕ�ûʳ��������Ч����Ӱ�죬�Դ�Ϊ����������������飬���ÿǾ��Ƕ�ûʳ����������ʺ���������Ϊ��Ӧָ�꣬����������Ҫ�������ҳ������Ƚϸߵ�������ϡ�
1.4 �������ûʳ���������������
�����������ºϳɿǾ�����ûʳ������˲���������25 ������ո���24 h���Ƶ����������100 mL����ˮ����ƿ�зֱ�����������ʹ��Ũ��Ϊ6��10��20��30��60��90��130 mg����������������ͬ�����£���0.2��1��3��10��24��48 h��ȡ����Һ����0.45 ��m��Ĥ��ⶨ����ȣ�����ûʳ���Ậ�������о��Ǿ�����ûʳ���������������ûʳ�����ͷŵ�Ӱ�졣
1.5 �Ǿ��ǡ�ûʳ������������2�ֵ�ˮ��������Ӱ��
����������пǾ�����ûʳ����ı���������m(�Ǿ���):m(ûʳ����):m(�����)Ϊ2:1:3��(1) ֱ��Ͷ����Ӧ�������ʷֱ�100 mL���ڶ����ڵ�ˮ������Һ��(Ҷ����a����Ϊ1 000 ��g/L����)��������������Ũ�ȷֱ�4��6��14��20��40��60��87 mg/L�ĿǾ��Ǵ����飬2��3��7��10��20��30��43 mg/L��ûʳ���ᴦ�����6��10��20��30��60��90��130 mg/L������������飬60 mg/L��ûʳ����Ϊ������������ʱ�����Ķ��գ��հ�����Ϊck��ÿ������3��ƽ������ÿ��1 d����һ��Ҷ����a�ĺ�����(2) ֱ��Ͷ����Ӧ�������ʷֱ�100 mL���ڶ����ڵĵ���С����Һ��(Ҷ����a����Ϊ1 000 ��g/L����)��������������Ũ�ȷֱ�Ϊ4��6��14��20��30��40��60 mg/L�ĿǾ��Ǵ����飬2��3��7��10��15��20��30��43 mg/L��ûʳ���ᴦ�����6��10��20��30��45��60��90��130 mg/L������������飬�հ�����Ϊck��ÿ������3��ƽ������ÿ��2 d����һ��Ҷ����a�ĺ�������������Ϊ��Ӧָ�꣬�۲������������ʱ��仯������Ч����Ӱ�졣
�������ҩЧ�ڣ�һ�㿼���ڼ���15 d����ϸ����������ʱ��ҩЧ��Ϊ���������������15 d����ϸ���ֿ�ʼ����ʱ����ҩЧ�ڽ϶�[15]�������۲�15 d�������ʵı仯��ȷ���������Ч�������Ч������
������������Ǿ�������Һ�г�Ϊ������������ͨ������������������������������Ҵ�ͳѪϸ�������������������Ϊ���ϴ�Ҷ������������й�����õ�����֮һ���غ������Ƿ��ܹ�˳��ͨ��������û����л����������ֳ��Ҷ����a����Ҷ���ص�һ�֣��京���ĸߵͿɷ�ӳ����ϸ�����й�����õ������Լ����������������������������ж�����Ҷ����a����ˣ�����ͨ���ⶨҶ����a�����������������ûʳ����ͿǾ��Ƕ�2�������Ӱ�졣Ҷ����a�������ø���ֲ�������(Phyto-PAM Phytoplankton Analyzer���¹�walz��˾)���вⶨ���ⶨ�����μ�����[16]��
1.6 ���ݴ���
(1) �Ǿ��Ƕ�ûʳ�������������ʽ����
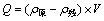 (1)
(1)
ʽ�У�QΪ��ûʳ�����������(mg)��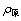 Ϊ��ʼûʳ��������Ũ��(mg/L)��
Ϊ��ʼûʳ��������Ũ��(mg/L)��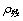 ��������Һ�в����ûʳ��������Ũ��(mg/L)��VΪûʳ������Һ���(mL)��
��������Һ�в����ûʳ��������Ũ��(mg/L)��VΪûʳ������Һ���(mL)��
(2) �Ǿ��Ƕ�ûʳ����������ȹ�ʽ���£�
 (2)
(2)
ʽ�У�PΪ�Ǿ��Ƕ�ûʳ�����������(mg/g)��mΪͶ�ӿǾ��ǵ�����(g)��
(3) �����ʵļ��㹫ʽΪ��
 (3)
(3)
ʽ�У�IΪ�����ʣ�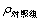 Ϊ������Ҷ���غ���(��g/L)��
Ϊ������Ҷ���غ���(��g/L)�� Ϊ������Ҷ���غ���(��g/L)��
Ϊ������Ҷ���غ���(��g/L)��
���ݲ���SPSS18.0����������ͳ�Ʒ���������Origin8.0������ͼ��Ҷ����a���������ظ��������������
2 ��������
2.1 �Ǿ�����ûʳ������������Ʊ�
�Ǿ�������ûʳ����ĵ�������������������ûʳ����Ũ��Ϊ����ʱ������������ûʳ����ʼŨ�ȵ����߶����ӣ������ٶ��ȿ���������ƽ���������ʱ仯��Χ�ϴ�����ûʳ��������Ũ��1 000 mg/Lʱ�ﵽ���ֵ85.76%�����Ǿ�������Ϊ����ʱ�������ȱ仯�ϴ����ſǾ������������Ӷ����ͣ��������ȿ�����������0.06 gʱȡ�����ֵ86.12%�����������͵�81.48%������������ڿǾ��ǹ��࣬���������ʹ�Ǿ�������Һ���Ӱ�����ɵģ��������¶�Ϊ����ʱ�������������¶ȵ����߶����ͣ���������¶�Ϊ25 �棻������ʱ��Ϊ����ʱ������������ʱ���ӳ������ӣ�ǰ30 min�������ʿ죬�������ټ�����
�������������1��ʾ���ڱ�ʵ�鷶Χ�ڣ��������Ⱥ�������Ϊ��Ӧָ�꣬���ȿ��������Ƚϴ�����3��4���ٿ��ǵ����3�������ʺܵ�ֻ��25.24%�������4�������ʽϸߣ�Ϊ51.94%�����ѡ�����4Ϊ���������ϡ��Ǿ�������ûʳ�������������Ϊ��20 mLûʳ������Һ�пǾ���������0.02 g���¶�Ϊ25 �棬����ʱ��Ϊ60 min��ûʳ�����ʼ����Ũ��Ϊ1 200 mg/L���ڸ������¿ɻ��623.095 mg/g�������ȡ�
��������Һ��ѭ��ʹ�ã�����������������Ʊ��ĿǾ�����ûʳ���������������Һ�в����ûʳ��������Ũ��Ϊ659.6 mg/L����ʱ��Һ���ԼΪ100 mL��������Һ���ܽ�54.04 mg��ûʳ�����ʹ��Һ����Ũ�ȴﵽ1 200 mg/L�������Ϊ�������ԭ�ϣ�ʹ��Һѭ�����á�����ûʳ������ˮ���ܽ�����������£����������������ʿ��ܻ���������Ӱ�졣������������Ʊ�ʱ����Ҫ��һ��ʵ����֤��Һ���ظ�ʹ�ô�����
��1 �Ǿ�������ûʳ����ʵ������������
Table 1 Orthogonal experimental results of adsorption capacity of Gallic acid by CTS

2.2 �������ûʳ�����������
��ͬ�������������ûʳ�������ͷź�Ũ�ȵ�Ӱ�죬��ͼ1��ʾ�����������Ũ�ȷֱ�Ϊ6��10��20��30��60��90��130 mg/Lʱ����Ͷ�ӳ����ͷ�Ũ���������ƽϿ죬�ͷ�����24 hʱ�ﵽ���ֵ���ֱ�Ϊ79.26%��90.80%��70.44%��84.34%��59.43%��48.46%��43.56%�����������Ͷ���������ӣ�ûʳ�����ͷ�Ũ�������ߣ��ͷ��ʳ������彵�͵����ƣ���˵����Һ���ܽ��ûʳ�����谭���������ûʳ������ͷţ���������������ﶯ̬ƽ��ʱ��ûʳ����Ũ���ȶ�������Һ��ûʳ����������֮����������ͷ�һ����ûʳ���ᣬ���ﵽ�µ�ƽ�⡣���⣬�����Ͷ����ˮ��֮�ḡ��ˮ�棬�ͷ�ʱ���㹻��Ͼ��ȵķֲ���������ɾֲ�Ũ�ȹ��ߡ�

ͼ1 �Ǿ�����ûʳ����������������Խ�����Ӱ��
Fig. 1 Effect of dosage of CTS-GA on desorption
2.3 �Ǿ��ǡ�ûʳ������������ˮ������������Ӱ��
�Ǿ��Ƕ�ˮ�������Ӱ����ͼ2(a)��(b)��ʾ���ɼ����������������û�б��ֳ���������(p��0.05)���Ǿ��dzʷ�ĩ״̬����������������û�о�����ʱ��������������ԿǾ��Ƕ�������������ñ��ֲ����ԡ���ͬ�����Ǿ��ǵ���������20%�ڲ��������ֳ��������������á�������Ǿ��Ƕ����������Ӱ�첻���ԣ�����������ԿǾ���û��Ͷ�����������ԡ�
ûʳ�����ˮ�������Ӱ����ͼ2(c)��(d)��ʾ���ɼ�����������ûʳ��������Ũ�ȵ����߶����ߡ���ûʳ��������Ũ��Ϊ2��3��7 mg/Lʱ������������Խ�������ûʳ��������Ũ��Ϊ10��20��30 mg/Lʱ����������ʱ����ӳ��������������г�Ч���壻��ûʳ��������Ũ��Ϊ43 mg/Lʱ��2 d�������ʿɴ�90%���ϣ����������۲�ĵ�16 d����������ά���ڽӽ�100%������16 d֮��Ҷ����a�������ֳ��������ƣ�˵��ûʳ�����ڸ�ʹ��Ũ���£���16 d�����ķֽ⣬Ũ�Ȳ����������������������ֳ�����ָ����������ƣ���ûʳ��������Ũ��Ϊ60 mg/L������ʱ����������24 dʱ��������ֳ��ָ��������ƣ���˵����Ũ�ȵ�Ͷ�����Բ��ʺϽ��г������塣
�������ˮ�����������Ч����ͼ2(e)��(f)��ʾ���ɼ�������Ч���������ʹ��Ũ�ȵ����߶����ߡ�6 mg/L��������������������ck��Ȳ����Բ�����(p��0.05)������Ũ�ȴ�����������(p��0.05)��������������������������Ũ��Ϊ10��20 mg/Lʱ�������������ߺͣ�������Ч��ɱ��ϸ���������������Ũ�ȴﵽ30 mg/Lʱ��2 d�������ʿɴ�90%���ϣ�5 d��ӽ�100%��������16 d�������Կɴ�90%���ϣ������˵�24 d�������½���80%���ڣ�˵��16 d�������ûʳ�����ͷŴ�������Һ�е�ûʳ���ᱻ�ֽ����ģ������������������ƣ������������Ũ��Ϊ60��90��130 mg/Lʱ��3 d�������ʿɴ�90%���ϣ�5 d��ɽӽ�100%�����۲�ĵ�24 d������������ά���ڽӽ�100%��˵���Ǿ�����ûʳ����������ڸ�ʹ��Ũ�������ڽϳ�ʱ���ڴﵽ�ϸߵ������ʡ�������Ϊ�Ǿ��Ƕ�ûʳ�����������ûʳ�������˱��������ö���ֹ����ͬʱ����������������һ�����ġ����Ͻ�����������������Ũ����30 mg/L����ʱ����ʱ��ɳ���15 d�������������Ũ����60 mg/L����ʱ����ʱ��ɳ���24 d��

ͼ2 CTS��GA��CTS-GA��ˮ������������Ӱ��
Fig. 2 Effects of CTS, Gallic acid and CTS-Gallic acid on growth of Microcystis flos-aquae
2.4 �Ǿ��ǡ�ûʳ�����������Ե���С����������Ӱ��
�Ǿ��ǶԵ���С�����Ӱ����ͼ3(a)��ʾ���Ǿ��ǶԵ���С���������Ӱ�첻����(p��0.05)��δ���������ö�����С�������Ҷ����a��������Ͷ��Ũ���ݶȿǾ��Ƕ�С�����Ӱ��û�б��ֳ����Ƶı仯���ƣ�˵��С����������ԿǾ��ǵ�Ͷ����û�������ԡ�ûʳ����Ե���С�����Ӱ����ͼ3(b)��ʾ��ûʳ�����ڵ�Ũ�ȵ�Ͷ������û�б��ֳ�������(p��0.05)�������ã���ûʳ��������Ũ��Ϊ43 mg/Lʱ��ǰ3 d��δ���ֳ����Ե����ƣ���������ûʳ����������������������������ɣ����������������ֳ����ӵ����ƣ���9 d�ﵽ���ֵΪ14%������������������ֱ����ģ���13 d��15 dʱ�����ʷֱ��½���12%��11%�������������ڿ���ǿ������ˮ������������ûʳ����Ũ�ȶԵ���С������������ò����ԡ�������Ե���С�����Ӱ����ͼ3(c)��ʾ����ͼ3�ɼ���������������������û�������Բ���(p��0.05)�������������Ũ��Ϊ60��90��130 mg/Lʱ�Ե���С�������������5%���ڣ�����3��Ũ��ȴ������������ˮ�������������˵����������ͷ�ûʳ�����γɵ�Ũ�Ȳ��������Ƶ���С������������Ե���С������ֳ���̬��ȫ�ԡ�

ͼ3 CTS��GA��CTS-GA�Ե���С����������Ӱ��
Fig. 3 Effects of CTS, Gallic acid and CTS-Gallic acidon growth of Chlorella pyrenoidosa
2.5 ��Ͷ�ӱ�������Ƚ϶�ˮ������������Ӱ��
���ղ�ͬ�ݶ����������Ӧ�����Ǿ��Ǻ�ûʳ��������ֱ���ͼ���бȽϣ���ͼ4��ʾ��ͼ4��ck�������տհף�y�����������k�����Ǿ��ǣ�m����ûʳ���ᡣ��ͼ4(a)�ɼ�����������ͷ�ûʳ�������ʽ����������ʱ�ֱ������Һ��Ͷ��ûʳ����Ҫ�ͣ����ͷ������������������������������ͼ4(b)�ɼ����������ֱ��Ͷ��ûʳ������ֳ��Ϻõ�����Ч�ʣ�����Ͷ�ӳ��ڣ�ֱ��Ͷ��ûʳ����ʹ��ͬʱ�����ķֽ⣬���Ժ��������ʽ��͡�����������ֻ����ͷţ����Խϳ�ʱ�������������������������Ͷ�����ϵͣ��ͷ����������������壻��ͼ4(c)�ɼ���ûʳ���������ڱ��ֳ��Ϻõ�����Ч��������֮����ͼ4(b)��ʾ��������ͬ��������Ũ���µ�ûʳ�������ȫ������������������������ֳ�����������Ч���������ͷ��ٶȽ����ͷ������Dz�������ȫ����������������ͼ4(d)�ɼ���ûʳ���������ڱ��ֳ��Ϻõ�����Ч�������ǵ��˵�10 d������ûʳ��������ģ������ʽ��ͣ�����ûʳ����ܳ�����������������������������������ǿ������Ч����12 d�ڼ�����Ҷ��������Ũ��ά����200 ��g/L���ڣ�����16 d֮������������ͷ��Լ����ģ������ֳ����������ƣ�ͼ4(e)��(f)��ʾûʳ���������Ч����ͼ4(d)��ʾ�����ƣ������нϺõ�����Ч�����ֱ��ڵ�5 d�͵�10 d֮�������ʱ�ͣ�����ָ�������������������ֳ�����������Ч������24 d�ڽ�Ҷ����a������Ũ��ά����25 ��g/L�ڣ����߳�ƽ�������ƣ������������ʱ��ɳ���24 d��
3 ����
3.1 ������пǾ��Ƕ�ûʳ������Ʊ��Լ�����
�Ǿ���δ���ܽ���Ա������ûʳ��������������Ҿ��нϸߵ�������������Ч�ʣ�������������Ʊ����̣��������������ڳ��µȼ������¾Ϳ��Խ��С������������25 ���£�������Ϊ1.5~2 a[17]�����������Ĵ��������Լ�ʹ�������ṩ�˱��ϡ�
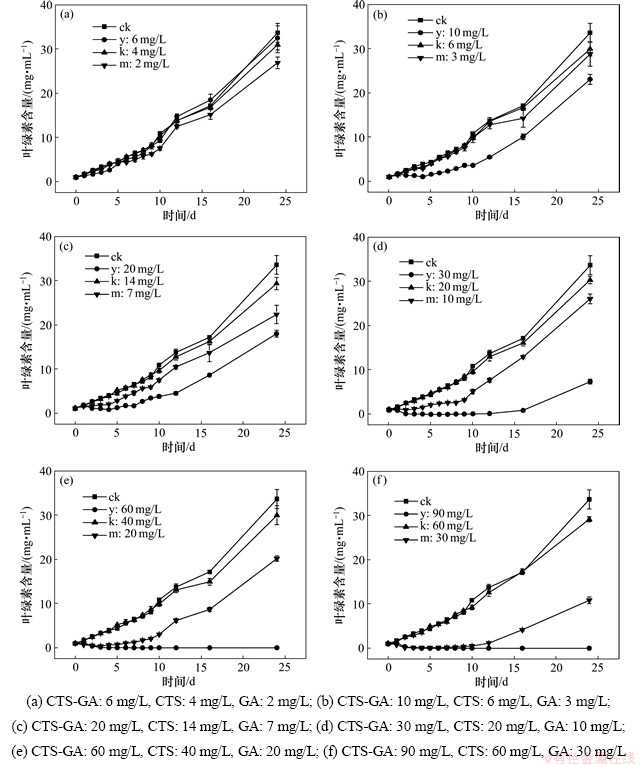
ͼ4 CTS-GA�Լ�GA��CTS��ˮ������Ҷ����a������Ӱ��
Fig. 4 Effects of CTS-GA, GA and CTS on Chlorophyll a content of Microcystis flos-aquae
������Ʊ�ʵ�����������Ǿ��Ƕ�ûʳ��������������ܴ�����������ûʳ����Ũ�ȵ����߶����ߡ���˵����һ�����Ǿ��Ƕ�ûʳ�������������������Һûʳ�����Ũ�ȣ����Ǿ���������ûʳ�����ϵ��ٶȵ���ûʳ����ӿǾ����Ͻ����������ٶ�ʱ���Ǿ��Ƕ�ûʳ����������ﵽƽ�⣬��ʱ���˱�ɵõ���Ӧ�����������������˵���Ǿ��Ƕ�ûʳ�����������һ���Է��Ĺ��̡��Ǿ��Ƿ����д��ڴ�����϶����ֱ������ûʳ���ᣬ�����������������ͷţ��������ͷ�ʵ����24 h��ɴﵽ�ϸߵ�Ũ�ȣ���������������������Ǿ��Ƿ����еĴ����ǻ��Ͱ���Ҳ������ûʳ�������ɻ�ѧ�������������������ͷ�[18]��������������Ի����ͷ�ûʳ������г������塣ûʳ������Ǿ��ǵĽ�ϲ�λ��ûʳ������������Ǿ��ǵİ���֮���γɵ������������ǻ�֮���γɵ��� ��[19]������ʵ������ʾ���Ǿ��Ƕ�ûʳ����������������ö���ֹ�䷢����������ͬʱ���������ģ��Ӷ��ﵽ�˳��������Ч���������ֱ��Ͷ��ûʳ�������ɵ�����ʱ�䲻������⡣
3.2 ��������ͷų���ûʳ������������
��ʵ���о����������ˮ������͵���С������������á��о����֣��������ˮ���������һ��������ЧӦ��������Ũ�ȵ���������Ч��Խ���ԡ�������ͷ�ûʳ���ᵽ���������ܱ���ϸ�������Ŀ������ʽ��⣬����Ϊ̼Դ����ϸ�����մ�л����������ʱ����ӳ��������������е�Ũ���Ʊػ����½��������͵�һ��Ũ��ʱ������Ч������ϸ����������ʹ�������½�[20-21]��������ṹ�о���ʾûʳ�����ϸ�������ò��Ƿ�����������еģ�����������ûʳ����ṹ�����е�������3�����ڵķ��ǻ�����Ҫ�Ļ��Ի���[22]����ˣ�ûʳ������н�ǿ�������ã���Ч���죬ͨ����һ��������ʵ�����̬��ȫ���ۣ����������ɳ�Ч���������
�����о�������ûʳ����ķ��ǻ����ڹ��ɽ���������Cu2+��Fe3+����ʱ���������������仯�����õ���Ҫ���ƣ�����һ����Ũ���£�����������������ô������������������������[23]����һ���о��������ڹ��ɽ������Ӵ���ʱ���������������յ�����H2O2������H2O2�ɽ�һ�����ϸ��֬�ʹ�����[24]��������ΪDZ�ڵĹ��������ͨ��������ԭѭ������ROS������Σ��ϸ��������[25]����������������䱽�����ǻ���λ�ú������йء���ʵ����õ�Bristol��BG11�������к���һ������Cu2+��Fe3+������Ȼˮ����һ��Ҳ����Cu2+��Fe3+�������������������������������������ûʳ����֮������ʱ������ƣ���Ũ����Һ�е�ûʳ���������������������������������ˮ��������и�ǿ���������á�����������ﱻ���ģ�Ũ�Ȳ����Զ���������������ã������������ʽ��͡�
3.3 �������2����������Ӱ��
ˮ��������������Ϊԭ�������ϸ������Ҫ�ɷ��ľ��ǣ�������Ҷ���塣����С������������Ϊ��������ϸ���ڼ�����Ҫ�ɷ�Ϊ��ά�أ�������Ҷ���塣�����У���Ũ��ûʳ�����ˮ���������������95%���ϣ����Ե���С�����������ȴ����10%������������������ϸ���ṹ�ϵIJ�ͬ����ɵ�Ӱ��Ч�����졣Zhu��[26]�о���ûʳ���������ͭ���������������������Ӱ�죬��������24 h��2.65 mg/L��ûʳ�������ͭ�������ϵͳII���������Ӵ�����������ż��٣��ֱ�Ϊ70.95%��40.77%(P��0.05)�����Ƕ����������û������Ч�������뱾�ĵ�ʵ����һ�£������������й�ϵͳ���Ƿ���������ʵ�һ������λ�㡣ͬʱ��ûʳ��������Ч������������ɽ������������ƻ���ϸ��Ĥ�ṹ��������ϸ�����������ѻ[27]����������ɽ�������ڼ��壬��ѧ����Ϊ���ǽ���ԭ���������������֮��ļ����������ûʳ����Բ�ͬϸ���ṹ����Ļ������û�����Ҫ��һ���о���
4 ����
(1) �Ǿ��Ƕ�ûʳ����������ܵ�ûʳ�����ʼŨ�ȡ��Ǿ����������¶��Լ�����ʱ������ص�Ӱ�죬�Ǿ�������ûʳ�������������Ϊ20 mL��ûʳ������Һ�пǾ�������Ϊ0.02 g���¶�Ϊ25 �棬����ʱ��Ϊ60 min��ûʳ�����ʼ����Ũ��Ϊ1 200 mg/L��
(2) ����������״����������������ͽ���ʱ���Ӱ�죬���ֳ��˻���Ч�������ҿǾ��Ƕ�ûʳ�������������ö���ֹ�併�⣬�ӳ��˵�����ûʳ�����ʹ��ʱ�䡣
(3) ���������ˮ���������Ч������30 mg/L���ϣ���������60 mg/L����ʱ����ʱ��ɳ���24 d�����ڳ�Ч���������ʵ��Ũ�ȷ�Χ�ڣ���������ڵ���С����û�б��ֳ���������Ч�������ֳ�������������̬��ȫ�ԡ�
�ο����ף�
[1] Klemas V. Remote sensing of algal blooms: An overview with case studies[J]. Journal of Coastal Research, 2012, 28(1A): 34-43.
[2] LI Zhaohui, WANG Qiang, RUAN Xiao, et al. Phenolics and plant allelopathy[J]. Molecules, 2010, 15(12): 8933-8952.
[3] John J, Sarada S. Role of phenolics in allelopathic interactions[J]. Allelopathy Journal, 2012, 29(2): 215-229.
[4] Mitrovic M, Jaric S, Djurdjevic L, et al. Allelopathic and Environmental implications of plant phenolic compounds[J]. Allelopathy Journal, 2012, 29(2): 177-197.
[5] Kamaya Y, Tsuboi S, Takada T, et al. Growth stimulation and inhibition effects of 4-hydroxybenzoic acid and some related compounds on the freshwater green alga Pseudokirchneriella subcapitata[J]. Archives of Environmental Contamination and Toxicology, 2006, 51(4): 537-541.
[6] Kovacik J, Klejdus B, Hedbavny J, et al. Effect of copper and salicylic acid on phenolic metabolites and free amino acids in Scenedesmus quadricauda(Chlorophyceae)[J]. Plant Science, 2010, 178(3): 307-311.
[7] Raman V, Ravi S. Effect of salicylic acid and methyl jasmonate on antioxidant systems of Haematococcus pluvialis[J]. Acta Physiologiae Plantarum, 2011, 33(3): 1043-1049.
[8] WANG Hongqiang, WU Zhengbin, ZHANG Shenghua. Relationship between the allelopathic activity and molecular structure of hydroxyl derivatives of benzoic acid and their effects on cyanobacterium Microcystis aeruginosa[J]. Allelopathy Journal, 2008, 22(1): 205-211.
[9] Pillai C K S, Paul W, Sharma C P. Chitin and chitosan polymers: Chemistry, solubility and fiber formation[J]. Progress In Polymer Science, 2009, 34(7): 641-678.
[10] ����, ��ƽ��, ����, ��. ������������ȥ������ೱ��[J]. �й�������ѧ, 2001, 121(1): 15-17.
LIANG Xiang, YIN Pinghe, ZHAO Ling, et al. Removing red tide algea in the sea by biomass carrier as algaecide[J]. China Environmental Science, 2001, 121(1): 15-17.
[11] �����, ��־��, �����. �Ǿ�����ͭ�������Chlorella pyrenoidosa��ȥ��[J]. ������ѧѧ��, 2011, 31(8): 1653-1659.
CHEN Yucheng, YANG Zhimin, LI Hongliang. Removal of chlorella pyrenoidosa by copper-chitosan algaecide[J]. Acta Scientiae Circumstantiae, 2011, 31(8): 1653-1659.
[12] Gerente C, Lee V K C, Le C P, et al. Application of chitosan for the removal of metals from wastewaters by adsorption: Mechanisms and models review[J]. Critical Reviews in Environmental Science and Technology, 2007, 37(1): 41-127.
[13] Ngah W S W, Teong L C, Hanafiah M A K M. Adsorption of dyes and heavy metal ions by chitosan composites: A review[J]. Carbohydrate Polymers, 2011, 83(4): 1446-1456.
[14] Ludovico P, Massimiliano F. Use of chitosan and chitosan- derivatives to remove arsenic from aqueous solutions[J]. Carbohydrate Research, 2012, 356: 86-92.
[15] ����, ����, ������. ��ͭ���ϳ�������ͭ������Ч���о�[J]. ����������ѧѧ��, 2009, 29(10): 910-913.
LI Xing, ZHAO Liang, YANG Yanling. A study on the inactivation of Microcystis Aeruginosa by manganese-copper composite algaecide[J]. Transactions of Beijing Institute of Technology, 2009, 29(10): 910-913.
[16] WANG Zhicong, LI Dunhai, QIN Hongjie, et al. An integrated method for removal of harmful cyanobacterial blooms in eutrophic lakes[J]. Environmental Pollution, 2012, 160(1): 34-41.
[17] ������, Ф��, ���ܾ�, ��. ûʳ��������ȶ����о�[J]. �ֲ���ѧ�빤ҵ, 2012, 32(4): 58-62.
GUO Manman, XIAO Zhuobing, PENG Mijun, et al. Thermal stability of gallic acid[J]. Chemistry and Industry of Forest Products, 2012, 32(4): 58-62.
[18] Cigdem Y, Gokce S K, Yesim A, et al. Gallic acid-loaded chitosan nanoparticles: A preliminary study[J]. European Biotechnology Congress, 2011, 22(1): 127-133.
[19] Wanvimol P, Garry R B, Suwabun C. Chitosan gallate as a novel potential polysaccharide antioxidant: An EPR study[J]. Carbohydrate Research, 2010, 345(1): 132-140.
[20] Poulson K L, Sieg R D, Prince E K, et al. Allelopathic compounds of a red tide dinoflagellate have species-specific and context-dependent impacts on phytoplankton[J]. Marine Ecology Progress Series, 2010, 416: 69-78.
[21] JIANG Dan, HUANG Lingfeng, LIN Shiquan, et al. Allelopathic effects of euhalophyte Salicornia bigelovii on marine alga Skeletonema costatum[J]. Allelopathy, 2010, 25(1): 163-172.
[22] Inoue M, Suzuki R, Sakaguchi N, et al. Selective induction of cell death in cancer cells by gallic acid[J]. Biological and Pharmaceutical Bulletin, 1995, 18(11): 1526-1530.
[23] Nakai S, Inoue Y, Hosomi M, et al. Myriophyllum spicatum- released allelopathic polyphenols inhibiting growth of blue-green algae Microcystis aeruginosa[J]. Water Research, 2000, 34(11): 3026-3032.
[24] Ayako F, Shinkji O, Mariko M, et al. (-)-Epigallocatechin gallate causes oxidative damage to isolated and cellular DNA[J]. Biochemical Phamacology, 2003, 66(9): 1769-1778.
[25] Wolf B, Christa M, Kurt S. Electron paramagnetic resonance studies of radical species of proanthocyanidins and gallate esters[J]. Archives of Biochemistry and Biophysics, 2000, 374(2): 347-355.
[26] ZHU Junying, LIU Biyun, WANG Jing, et al. Study on the mechanism of allelopathic influence on cyanobacteria and chlorophytes by submerged macrophyte (Myriophyllum spicatum) and its secretion[J]. Aquatic Toxicology, 2010, 98(2): 196-203.
[27] ��ά��, ������, ������. ����������ʶ���������ɽ����������Ӱ��[J]. �й�������ѧ, 2005, 25(4): 417-419.
YANG Weidong, ZHANG Xinlian, LIU Jiesheng. The influence of allelochemicalo on the growth of Alexandrium tamarense[J]. China Environmental Science, 2005, 25(4): 417-419.
(�༭ ����ƽ)
�ո����ڣ�2013-07-01�������ڣ�2014-03-14
������Ŀ��������Ȼ��ѧ����������Ŀ(20777021)����������ѧ�����о��ص���Ŀ(210253)������ʡ��Ȼ��ѧ����������Ŀ(2010J01043, D0610012)
ͨ�����ߣ�����ӿ(1969-)���У��ӱ�ʯ��ׯ�ˣ���ʿ�����ڣ�����ˮ��Ⱦ���μ�������Դ�����о����绰��18005020185��E-mail: guopeiyong@sina.com
ժҪ���������������о��Ǿ���(CTS)����ûʳ����(GA)��Ӱ�����أ������ó��Ǿ��Ƕ�ûʳ�����������������ǣ�ûʳ������Һ��ʼ����Ũ��Ϊ1 200 mg/L��20 mLûʳ������Һ�пǾ�������Ϊ0.02 g����ʱ��Ϊ60 min���¶�Ϊ25 �档�ڴ��������Ʊ���ûʳ������Ǿ��ǵ�������Ϊ623 mg/g������������о�������ͷ��ʡ�Ȼ��̽�ָ��������ˮ������(Microcystis flos-aquae)�͵���С����(Chlorella pyrenoidosa)������Ӱ�졣�о�������������������������ˮ����������������������ã������������Ũ��Ϊ30 mg/Lʱ������ʱ�佫�ﵽ15 d�������������Ũ��Ϊ60��90��130 mg/Lʱ������ʱ�佫����24 d�������ʸߴ�99%��������15 d�ڣ��������δ�Ե���С������ֳ����Ե��������ã������������Ũ��Ϊ60 mg/Lʱ�������ʽ�Ϊ5%����ˣ��Ǿ�����ûʳ�������������������Ч����ˮ������ˮ��������������Ե���С�����Ӱ�������
