
Cu2+��Zn2+��������ķ��롢�������主������
���ƹ����� ������ �ȣ����峬�������壬��˼��
(���ϴ�ѧ ������ѧ�빤��ѧԺ������ ��ɳ��410082)
ժ Ҫ��
ժ Ҫ���Ӻ���ʡ����������ͭпβɰ�������ڷ��롢�����õ�1�꿹ͭ��п�ľ���CTB430-1�������þ�����̬�Ͳⶨ����ITS�������м������֣��о���ͬpHֵ���������ӳ�ʼŨ�Ⱥͽ������ӹ���Ծ��������������ؽ�����Ӱ�죻�ֱ���ÿ���˹�������͵������ⶨ�����ڿ����Ե��ͻ�ԭ�Թ����ĺ������о���������������þ���ΪAspergillus flavus�����Cu2+��Zn2+������־�����Ũ�ȷֱ�Ϊ400��800 mg/L�������������������ҺpHֵ���������ӳ�ʼ����Ũ�Ⱥͽ������ӹ����Ӱ�죬Cu2+��Zn2+�ĸ������ֱ���������Ũ��Ϊ200��250 mg/Lʱ�ﵽ���Ϊ30.82��40.37 mg/g�������ڿ����Ե��ͻ�ԭ�Թ����ĺ��������仯�����������Zn2+���н�ǿ����Ӧ�����Ϳ��ԣ���ԭ�Թ������ܻ���Cu2+��Zn2+��CTB430-1���������ˡ�
�ؼ��ʣ�
�����ͭ��п������������������������ԭ�Թ�������
��ͼ����ţ�X51 ���ױ�ʶ�룺A ���±�ţ�1672-7207(2009)01-0060-07
Isolation, identification and its bioaccumulation characteristics of
a fungus strain with resistance to Cu2+ and Zn2+
LIU Yun-guo, FAN Ting, ZHOU Na, HE Yi-chao, Min Zhong-yi, WU Si-dan
(College of Environmental Science and Engineering, Hunan University, Changsha 410082, China)
Abstract: A fungus CTB430-1 with resistance to copper and zinc was isolated from soil at a copper and Zinc Tailing of Linxiang in Yueyang city, Hunan province. The fungus was identified by analyzing the morphology and measuring the ITS sequence. The effects of pH, initial concentration, co-ions on the growth of CTB430-1 and bioaccumulation of Cu2+ and Zn2+ were studied. The protein and glutathione in the cells were determined by using the coomassie brilliant blue method and iodometric method, respectively. The results show that the strain is identified as Aspergillus flavus. Its minimal inhibitory concentration for Cu2+ and Zn2+are 400 and 800 mg/L, respectively. pH, initial concentration, co-ions in solution can affect the growth of CTB430-1 and bioaccumulation of Cu2+ and Zn2+, and the maximum bioaccumulation capacities of Cu2+ and Zn2+ are 30.82 and 40.37 mg/g at initial concentrations of 200 and 250 mg/L, respectively. Furthermore, the changes of protein and glutathione in the cells show that the strain holds high adaptive abilities and tolerance of zinc, and glutathione can alleviate the oxidation stress of copper and zinc.
Key words: fungus; copper; zinc; resistance; bioaccumulation; protein; glutathione
�ؽ�����Ⱦ����Ȼ����ͨ���Ծ����û����オ��ȥ����������Ȼ�����еĹ��ޡ�Ǩ�ơ�ת���Լ��������ۻ�ЧӦ�����⣬�ѳ�Ϊ������ѧ�о����ȵ�[1-2]��Ŀǰ����������Ҫ�û�ѧ��������������ԭ�������ӽ������������ؽ����ķ�ˮ���ɱ��ϸ�[3-4]���������������ǿ�������Ե�Ũ�Ⱥ��ؽ�����ˮ����������Ч���ã��ɱ��ͣ�������й�����Ӧ��ǰ ��[5-7]���ڴˣ��������߶�Cu2+��Zn2+���и�Ч���Եĸ�������CTB430-1��ͨ������������̬�ͲⶨITS�������м������֡�ͬʱ���о������Cu2+��Zn2+�Ŀ��Ժ������ԡ��ؽ��������¾����ڿ����Ե��ͻ�ԭ�Թ����ĵĺ����仯��̽��������ؽ����Ŀ��Ի�����
1 ʵ����Ϻͷ���
1.1 ʵ�����
a. ������������(ɸѡ������)�������� 10 g��������5 g��KH2PO4 1 g��MgSO4��7H2O 0.5 g����֬15~ 20 g������ˮ1 L��Ϊ����Ч����ϸ���ͷ��߾�����������1%�ϼ�����ˮ��Һ3.3 mL��1%��ùϡ��Һ3 mL��
b. ����������(����������)��ȥƤ������500 g (����ϴ��ȥƤ����ˮ����������15 min)��������20 g����֬20 g������ˮ1 L��
c. ����������(Һ��������)��������20 g��������10 g��NaCl 0.2 g��CaCl2 0.1 g��KCl 0.1 g��K2HPO4 0.5 g��NaHCO3 0.05 g��MgSO4 0.25 g��FeSO4��7H2O 5 mg������ˮ1 L��
1.2 ����ɸѡ
�Ӻ���ʡ����������ͭпβɰ��ȡ��������10 g��������װ�뺬��90 mL��ˮ������ƿ�У��ò������ɢ�����ȣ��Ƴ�������Һ����Ϳ����Ϳ���������Ϲ���������ƽ���ϣ���28 ������5~7 d��Ȼ����ѡ����������������������߷��룬�������������ҳ�������״����ɫ��ͬ�ľ��꣬�����ƶ�Ϊͬһ���֡���ɸѡ���ľ����������������б���У���28 ������5 d������4 ��ı������б��汸�á�
�������ľ��䡢ϸ����̬�����ж�������[8]�з������С�
1.3 ITS�������вⶨ�ͼ���
�Ծ������ ITS����������ȡ������������10 ��L�ѽ�Һ�У���80 �����15 min��ȡ1 ��L��Ϊģ�棬ʹ��TaKaRa Fungi Indentification PCR Kit(Code No.D317)PCR����Ŀ��Ƭ�Σ���ITS1 Forword primer/ITS4 Reverse primerΪ���������������ITS�������У������ɱ����﹤��(����)����˾��ɡ�
���ⶨ�������ύ��NCBI(�����������\����Ϣ����)���ݿ⣬Ӧ��BLAST���������ݿ������еĻ������н���ͬԴ�ԱȽϷ�����
1.4 ����Һ�Ʊ�
���������£�ȡ����5 mg����װ��200 mL��ˮ��500 mL��ƿ�У����¶�Ϊ28 �桢ת��Ϊ120 r/min��������2 h����4 ��ı������б��汸�á�
1.5 ����Ŀ���ʵ��
��1 mL����Һ����Cu2+��Zn2+����Ũ�Ȳ�ͬ(0~1 g/L)��Һ��������100 mL�У���28 �桢ת��Ϊ120 r/min������������7 d�����ܶȷ��ⶨ���ֵ�����������������Ŀ��ԡ�
1.6 ����ĸ�������
1.6.1 pHֵ�Ծ���CTB430-1����������Cu2+��Zn2+��Ӱ��
��1 mL����Һ����װ��100 mLҺ��������(Cu2+��Zn2+����Ũ�Ⱦ�Ϊ50 mg/L)����ƿ�У����ʼ��pHֵ��ΧΪ2~6����28 �桢ת��Ϊ120 r/min��������������7 d��
1.6.2 ��ͬŨ���ؽ����Ծ���CTB430-1����������Cu2+��Zn2+��Ӱ��
��1 mL����Һ����װ��100 mLҺ������������ƿ�У�Cu2+��Zn2+����Ũ�ȷ�ΧΪ25~200 mg/L����28 �桢ת��Ϊ120 r/minʱ������7 d��
1.6.3 Cu2+��Zn2+����ʱ�Ծ���CTB430-1����������Cu2+��Zn2+��Ӱ��
��1 mL����Һ����װ��100 mLҺ������������ƿ�У����벻ͬ����Ũ�ȵ�Cu2+��Zn2+(0��25��50��100 mg/L)����28 �桢ת��Ϊ120 r/minʱ������7 d��
1.7 ���꿹�Ի�������
��1 mL����Һ����װ��100 mLҺ������������ƿ�У�Cu2+��Zn2+����Ũ�ȷ�Χ�ֱ�Ϊ0~200 mg/L����28 �桢ת��Ϊ20 r/minʱ������7 d���ⶨ��ͬ�ؽ���Ũ���¾������ڿ����Ե��ͻ�ԭ�Թ����ĵĺ����仯��
����ÿ��ʵ�����3�������ȡƽ��ֵ��
1.8 �ⶨ����
1.8.1 �����������ⶨ
ʵ����ɺ����˵ľ���������ˮ��ϴ�ɾ������¶�Ϊ60 ��ĺ����к��������㶨���ⶨ������������
1.8.2 ����Cu2+��Zn2+������
ʵ����ɺ�ÿ����Ʒȡ1 mL������Һ�ⶨ�ؽ�����Ũ�ȣ������ؽ����ij�ʼŨ�ȼ��������ؽ����ĸ�������

1.8.3 �ؽ���Ũ�Ȳⶨ
��ԭ�����շֹ��ȼ�(PE AA700 U.S.A.)�ⶨ��Ӧ��ʣ����Һ���ؽ���Ũ�ȡ�
1.8.4 �����Ե�����ȡ�Ͳⶨ
ȡ1.0 g����ϴ�ɾ������ʾ��壬����5.0 mL Tris-HCL(pH7.0)��ȡҺ��Һ����ĥ����ת��Ϊ5 000 r/min���¶�Ϊ4 �������������30 min���õ�������Һ�ÿ���˹�����������Լ���(�Ͼ��������﹤���о�������)������������ֹ��ȼƲⶨ�����Ե���Ũ�ȡ�
1.8.5 ��ԭ�Թ����ĵ���ȡ�Ͳⶨ
ȡ1 g��ɵľ����ĩ������˳�����4 mL����ˮ��5 mL��ˮ����ֻ�Ϻ����95~100 ��ˮԡ����ˮԡ10 min��Ȼ���������ڱ�ˮ�����䣬��ת��Ϊ5 000 r/minʱ����10 min��ȡ��������Һ���õ��� ��[9]�ⶨ��ԭ�Թ����ĵĺ�����
2 ���������
2.1 �����ɸѡ
����η���ɸѡ���õ�1���Cu2+��Zn2+���п��Եľ��꣬����ΪCTB430-1�������������������о���η��뻮�ߣ���28 ������72 h�����겼������ƽ����˿���ɣ�����ʻ���ɫ�������Գʺ�ɫ���������۲죬��˿���֦�����и�Ĥ���д����ķ������ӹ��������������ε����ң������ұ����������ʷ���״���е�С����ÿ��С�����˲����ɴ��ķ������ӣ��ʻ���ɫ�����ݾ��䡢ϸ����̬�����������ж�Ϊ��ù��(Aspergillus)��
2.2 ITS�������вⶨ�����
���ⶨ��CTB430-1��ITS��������(GeneBank�ϵ�ע���Ϊ FJ389893)��NCBI�Ͻ���BLAST�Ƚϣ��������������Aspergillus����Aspergillus flavus�ȶ���ֵ�ITS�������о���100%��ͬԴ�ԣ���Aspergillus oryzae�ȶ���־���99%��ͬԴ�ԣ���ˣ������þ���ΪAspergillus flavus��
2.3 ����CTB430-1���ؽ���Cu2+��Zn2+�Ŀ���
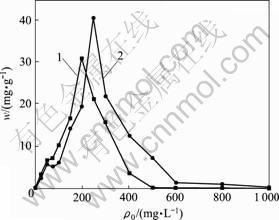
Cu2+��Zn2+��CTB430-1����������Ӱ��ֱ��ͼ1��ͼ2(���У�mΪ���帻������g��wΪ������������mg/g)����ͼ1��ͼ2��֪������CTB430-1��Cu2+��Zn2+�����п��ԣ���Cu2+��Zn2+����Ũ�ȷֱ����50��100 mg/Lʱ���Ծ���CTB430-1���������ò����ԡ�Cu2+��Zn2+��CTB430-1���������־�����Ũ�ȷֱ�Ϊ400��800 mg/L��Trichoderma atroviride[10]��Cu2+��Zn2+����Ũ�ȷֱ�Ϊ400��800mg/Lʱ���ܴ�Pseudomonas aeruginosa Asu 6a[11]��Cu2+��Zn2+����־�����Ũ�ȷֱ�Ϊ403.2��598 mg/L���ɼ��� CTB430-1�����Cu2+��Zn2+���н�ǿ�Ŀ��ԡ�

1��Cu2+��2��Zn2+
ͼ1 Cu2+��Zn2+��CTB430-1������Ӱ��
Fig.1 Effects of Cu2+ and Zn2+ concentration on growth of CTB430-1
1��Cu2+��2��Zn2+
ͼ2 CTB430-1��Cu2+��Zn2+�ĸ�����Ӱ��
Fig.2 Effects of CTB430-1 on accumulation of Cu2+ and Zn2+
����Cu2+��Zn2+��ʼ����Ũ�ȵ����ӣ�����CTB430-1�����������ͣ�����Խ������ӵĸ����������Ӻ���٣�Cu2+��Zn2+�ĸ������ֱ���������Ũ��Ϊ200��250 mg/Lʱ�ﵽ���Ϊ30.82��40.37 mg/g��ϸ������Ĥ�ı��渻��������ֿ��ؽ������Ӷ��Ե��ֶ�֮һ���ڽϸ�Ũ�ȵ��ؽ������ӻ����У������������һ�������ؽ������ӣ��̼����ڿ��Ի������У��ٽ����������ؽ������õ�ø���ɣ�ͬʱ��ͨ����Ĥ�ɷֵĸı�ٽ��ؽ������Ӹ�������Ũ�ȵ�Cu2+��Zn2+�Ծ������һ���Ķ��ԣ����¾����³´�л���Ժ����������͡�
2.4 ����CTB430-1���������ؽ����ĸ�������
2.4.1 pHֵ�Ծ���CTB430-1����������Cu2+��Zn2+��Ӱ��
pHֵ�Ծ���CTB430-1����������Cu2+��Zn2+��Ӱ��ֱ��ͼ3��ͼ4����ͼ3��ͼ4��֪���� pH��5ʱ�����������������������Խ�ǿ����ԭ����Ҫ�Ǵ��������ͨ�����������Ժ������Ի�����pHֵ���ͽ����ƾ���Ļ��Ժ�������pHֵ�ϵ�ʱ������Һ�д���H+��������Ӿ��������ڻ���λ������ƾ���Խ������ӵĸ���������������������Ŵ�����ɣ������ڽ������ӵĸ�������pHֵ�ϵ�ʱ�书���Ŵ�����ɣ�����������ӷ�������[12]��pHֵΪ5ʱ��������Cu2+��Zn2+��Һ���������������ﵽ��ߣ����������ֱ�Ϊ0.66��0.80 g���������ֱ�Ϊ5.31��6.07 mg/g��
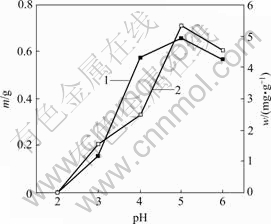
1������������w��2�����帻����m
ͼ3 Cu2+����Ũ��Ϊ50 mg/LʱpHֵ�Ծ���CTB430-1����������Cu2+��Ӱ��
Fig.3 Effects of pH value on growth of CTB430-1 and accumulation of Cu2+ when concentration of Cu2+ is 50 mg/L
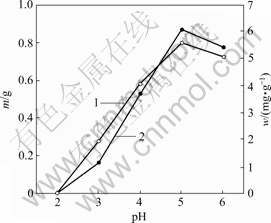
1������������w��2�����帻����m
ͼ4 Zn2+����Ũ��Ϊ50 mg/LʱpHֵ�Ծ���CTB430-1����������Zn2+��Ӱ��
Fig.4 Effects of pH value on growth of CTB430-1 and accumulation of Zn2+ when concentration of Zn2+ is 50 mg/L
2.4.2 �ؽ�����ͬ��ʼŨ�ȶԾ���CTB430-1����������Cu2+��Zn2+��Ӱ��
�ؽ������ӳ�ʼŨ�ȶԾ���CTB430-1��������Cu2+��Zn2+��Ӱ��ֱ��ͼ5��ͼ6����ͼ5��ͼ6��֪��Cu2+��Zn2+Ũ�����ӶԾ���������DZ�ڵ��������ã����¾����������������ӣ�����������������˵�����긻����Ҫ�������������Ĵ�л�����[13]��Cu2+��Zn2+�������ھ���������ʼ�νϵͣ�����ʱ���ӳ������������������������ӣ��ھ����������ﵽƽ��ʱ������������Cu2+��Zn2+�������ֱ���4 d��5 dʱ�ﵽƽ�⡣�ɼ���Cu2+�Ծ��������������ý�Zn2+�Ĵ�
1��Cu2+ 25 mg/L; 2��Cu2+ 50 mg/L; 3��Cu2+ 100 mg/L;
4��Zn2+ 25 mg/L; 5��Zn2+ 50 mg/L; 6��Zn2+ 100 mg/L
ͼ5 �ؽ������ӳ�ʼŨ�ȶԾ���CTB430-1������Ӱ��
Fig.5 Effects of initial concentration of Cu2+and Zn2+on growth of CTB430-1

1��Cu2+ 25 mg/L; 2��Cu2+ 50 mg/L; 3��Cu2+ 100 mg/L;
4��Zn2+ 25 mg/L; 5��Zn2+ 50 mg/L; 6��Zn2+ 100 mg/L
ͼ6 �ؽ������ӳ�ʼŨ�ȶԾ���CTB430-1����Cu2+��Zn2+��Ӱ��
Fig.6 Effects of initial concentration on accumulation of Cu2+and Zn2+ of CTB430-1
2.4.3 Cu2+��Zn2+����ʱ�Ծ���CTB430-1������������Ӱ��
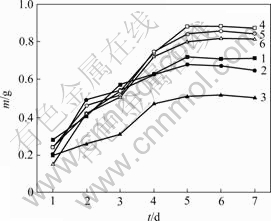
Cu2+��Zn2+����ʱ�Ծ���CTB430-1������������Ӱ���ͼ7����ͼ7��֪��Cu2+��Zn2+����ʱ�Ծ���CTB430-1�����������ô��ڵ������Ӵ��ڵ����ã���������������2�ֽ���Ũ�����ӵݼ��������Cu2+��Zn2+�ĸ��������ֱ���(100��50) mg/L��(25��100) mg/Lʱ�ﵽ���Ϊ7.65��14.94 mg/g����˵���ڹ���ϵͳ�ڣ�����������Ũ�Ƚϸ�ʱ���侺��������ǿ������Cu2+��Zn2+����ʱ����Ũ��Cu2+��Zn2+�ܴٽ�����Ը�Ũ��Zn2+��Cu2+����ĸ��������߹���Ũ�ȶ��ϸ�ʱ�����긻�������½����ɼ������긻��Zn2+�����ܵ����������ý�С����Cd2+��Zn2+��Pb2+����ʱ��Penicilium simplicissimum[14]�����������ӵ�������С�ڵ����Ӵ���ʱ��������������Zn2+�����������ܵ�������������С���뱾�о��������һ���������ԡ�
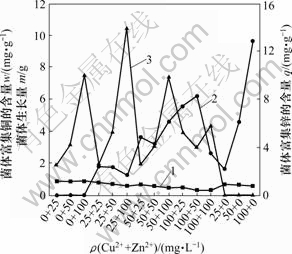
1��������������2�����帻��ͭ�ĺ�����
3�����帻��п�ĺ���
ͼ7 Cu2+��Zn2+��ͬ����ʱ�Ծ���CTB430-1������������Ӱ��
Fig.7 Effects of co-ions on growth of CTB430-1 and accumulation of Cu2+and Zn2+
2.5 ����CTB430-1���ؽ����Ŀ��Ի���
2.5.1 ��ͬŨ���ؽ����Ծ���CTB430-1���ڿ����Ե�������Ӱ��
�о�������ϸ������Ӧ����в�ȵ�������������Ҫ���������ʵĺϳ��뽵�⣬����Ӧ�µĻ���[15]���ؽ����Ծ���CTB430-1���ڿ����Ե�������Ӱ���ͼ8����ͼ8��֪����Cu2+��Zn2+�����¿����Ե��������������½������������ƣ����������½�����Cu2+��Zn2+Ũ�Ƚϵ�ʱ�����Ե������������ͣ���ԭ�������Cu2+��Zn2+�����˿����Ե��ϳɲ��յ������ʵĽ��⣬�����˿����Ե������������ؽ���Ũ�����ӣ������Ե��������ӣ�����������澳в�ȵ�һ������������Ӧ����ԭ�������ϸ���ṹ���������ƻ�ʱ��Ӧ����Ӧ����һ���棬�������澳�µ���Ӧ���ּ�������Ӧ������Ϊ����������Կ��Ե�ָ�ꡣ����Cu2+��Zn2+Ũ�ȼ������ӣ���CTB430-1�Ķ���Ҳ���������Ե�������ʼ��С��ϸ����л�ٶȼ�������Cu2+��Zn2+в�ȵ���Ӧ����ǿ��Heyser��[16]��Ϊ���������Ϳ�ʹϸ����л�ٶȼ������Ӷ���ǿ����ϸ���Ի���в�ȵ���Ӧ�������뱾ʵ������һ�¡�

1��Cu2+; 2��Zn2+
ͼ8 �ؽ������ӳ�ʼŨ�ȶԾ���CTB430-1���ڿ����Ե�������Ӱ��
Fig.8 Effects of initial concentration of Cu2+and Zn2+on soluble protein contents in cells of CTB430-1
�ܵ���˵������CTB430-1���ڿ����Ե������仯������Ҫ�������Cu2+��Zn2+���п��ԣ��Ҷ�Zn2+���Խ�ǿ��
2.5.2 ��ͬ�ؽ���Ũ�ȶԾ���CTB430-1���ڻ�ԭ�Թ����ĵ�Ӱ��
�ؽ������ӳ�ʼ����Ũ�ȶԾ���CTB430-1���ڻ�ԭ�Թ����ĺ�����Ӱ���ͼ9����ͼ9��֪������Cu2+����Ũ�ȵ����ӣ�����CTB430-1���ڻ�ԭ�Թ�����(GSH)�ĺ��������ߣ�Ȼ�����½�������Zn2+����Ũ�����ӣ�GSH�ĺ��������Ӻͣ���50~150 mg/L���ֳ��������Ӻ����ơ��ñ仯���ؽ��������ʺ�Ũ���йء����⣬�������Ӵ���ʱ��������GSH���������ڿհ�ʱ�ĺ�����������������Դ̼���������GSH���������ؽ�����ϸ�����������ˡ�GSH���������ڷǵ�������Ҫ��Դ�������ϻ��������ؽ���ֱ�ӽ��[17]��ͬʱ��GSH������������Ҫ�Ŀ��������ʣ������ϸ���ڻ����������������� ��[18]����Cu2+��Zn2+����Ũ��Ϊ25 mg/Lʱ��GSH�����ﵽ��������ڻ��������ʴ������ӣ�����Cu2+��Zn2+����Ũ�ȴ���25 mg/Lʱ��GSH�������͡���ԭ������Ǿ������ڿ�����ϵͳ�����ɷֵĽ��룬�����˻�����Ũ�ȣ����߾�������GSH��������ӷ�����Ӧ������������ӵĶ��ԡ����⣬��Ũ�Ƚ������ӶԾ���������˶�����GSH�IJ���[19]��
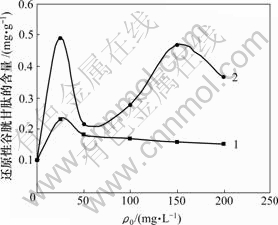
1��Cu2+; 2��Zn2+
ͼ9 �ؽ������ӳ�ʼŨ�ȶԾ���CTB430-1���ڻ�ԭ�Թ����ĺ�����Ӱ��
Fig.9 Effects of initial concentration of Cu2+and Zn2+on glutathione contents in cells of CTB430-1
3 �� ��
a. �Ӻ���ʡ����������ͭпβɰ��������ɸѡ�õ�1���Cu2+��Zn2+���и�Ч���Եľ���CTB430-1��ͨ��������̬������ITS�������вⶨ�����������þ���Ϊ����ù(Aspergillus flavus)��
b. Cu2+��Zn2+�Ծ���CTB430-1������־�Ũ�ȷֱ�Ϊ400��800 mg/L��Cu2+��Zn2+�ĸ������ֱ���������Ũ��Ϊ200��250 mg/Lʱ�ﵽ���Ϊ30.82��40.37 mg/g���������������������ӵ���������Һ��ʼpHֵ���������ӳ�ʼ����Ũ���Լ��������ӹ����Ӱ�졣��pH��5ʱ�����������������������Խ�ǿ������Cu2+��Zn2+����Ũ�����ӣ��Ծ�����������������������ȶ��ԣ�Cu2+���������ýϴ�Cu2+��Zn2+����ʱ�Ծ���CTB430-1�����������ô��ڵ������Ӵ���ʱ�����ã����߹�������Ũ�ȶ��ϸ�ʱ������CTB430-1���������ϵ͡�
c. ��Cu2+��Zn2+��ͬŨ�������£������ڿ����Ե��ͻ�ԭ�Թ����ĺ����仯����˵�����þ����Zn2+���н�ǿ�Ŀ��ԣ���ԭ�Թ������ܻ���Cu2+��Zn2+��CTB430-1���������ˡ�
�ο����ף�
[1] Artola A, Balaguer M D, Rigola M. Heavy metal binding to anaerobic sludge[J]. Water Research, 1997, 31(5): 997-1004.
[2] Malik A. Metal bioremediation through growing cells[J]. Environmental International, 2004, 30(2): 261-278.
[3] Pagnanelli F, Trifoni M, Beolchini F, et al. Equilibrium biosorption studies in single and multi-metal systems[J]. Process Biochemistry, 2001, 37(2): 115-124.
[4] Kadirvelu K, Senthilkumar P, Thamaraiselvi K, et al. Activated carbon prepared from biomass as adsorbent: elimination of Ni(��) from aqueous solution[J]. Bioresoure Technology, 2002, 81(1): 87-90.
[5] Lodeiro P, Barriada J L, Herrero R, et al. The marine macroalga Cystoseira baccata as biosorbent for cadmium(��) and lead(��) removal: Kinetic and equilibrium studies[J]. Environmental Pollution, 2006,142(2): 264-273.
[6] LIU Yun-guo, FAN Ting, ZENG Gung-ming, et al. Removal of cadmium and zinc ions from aqueous solution by living Aspergillus niger[J]. Transactions of Nonferrous Metals Society of China, 2006, 16(3): 681-686.
[7] Shankar C, Sridevi D, Joonhong P, et al. Biosorption of chromium and nickel by heavy metal resistant fungal and bacterial isolates[J]. Journal of Hazardous Materials, 2007, 146(1/2): 270-277.
[8] κ����. ��������ֲ�[M]. �Ϻ�: �Ϻ���ѧ����������, 1979: 129-136.
WEI Jing-chao. Fungi identification handbook[M]. Shanghai: Shanghai Science and Technology Press, 1979: 129-136.
[9] ���ͻ�. ��ԭ�Թ�������ȡ������̽[J]. ������ѧԺѧ��, 2003, 12(2): 49-52.
AN Xian-hui. A preliminary study on the extracting method of glutathione[J]. Journal of Huaihai Institute of Technology, 2003, 12(2): 49-52.
[10] L��pez E E, V��zquez C. Tolerance and uptake of heavy metals by Trichoderma atroviride isolated from sludge[J]. Chemosphere, 2003, 50(1): 137-143.
[11] Hassan S H A, Abskharon R N N, Gad El-Rab S M F, et al. Isolation, characterization of heavy metal resistant strain of Pseudomonas aeruginosa isolated from polluted sites in Assiut city, Egypt[J]. Journal of Basic Microbiology, 2008, 48(3): 168-176.
[12] Yan G, Viraraghavan T. Heavy metal removal from aqueous solution by fungus Mucor rouxii[J]. Water Research, 2003, 37(18): 4486-4496.
[13] Uslu G, Dursun A Y, Ekiz H I, et al. The effect of Cd(��), Pb(��) and Cu(��) ions on the growth and bioaccumulation properties of Rhizopus arrhizus[J].Process Biochemistry, 2003, 39(1): 105-110.
[14] Ting F, Yun-Guo L, Bao-ying F, et al. Biosorption of cadmium(��), zinc(��) and lead(��) by Penicillium simplicissimum: Isotherms, kinetics and thermodynamics[J]. Journal of Hazardous Materials, 2008, 160(2/3): 655-661.
[15] Banerjee B D, SETH V, Bahattacharya A. Biochemical effects of some pesticides on lipid peroxidation and free-radical scavengers[J]. Toxicology Letters, 1999, 107(1/3): 33-47.
[16] Heyser J W, Nabors M W. Growth, water content and solute accumulation of two tobacco cell lines cultured on sodium chloride, dextran, and polyethylene glycol[J]. Plant Physiology, 1981, 68: 1454-1459.
[17] ���ı�, ��ʤ. ������ؽ�����������ⶾ����[J]. ������ѧ, 2004, 16(3): 265-272.
ZHOU Wen-bin, QIU Bao-sheng. Mechanisms for heavy metal detoxification and tolerance in algae[J]. Journal of Lake Sciences, 2004, 16(3): 265-272.
[18] Ana I G L, Sofia C C, Etelvina M A P F. Glutathione-mediated cadmium sequestration in Rhizobium leguminosarum[J]. Enzyme and Microbial Technology, 2006, 39(4): 763-769.
[19] Saint D, Labrot F, Narbonne J F, et al. Glutathione, glutathione-related enzymes, and catalase activities in the earthworm Eisenia fetida andrei[J]. Archives of Environmental Contamination and Toxicology, 1998, 35(4): 602-614.
�ո����ڣ�2008-11-10�������ڣ�2009-01-05
������Ŀ����ʮһ�塱���ҿƼ�֧�żƻ���Ŀ(2006BAD03A1704��2006BAD03A1706)
ͨ�����ߣ����ƹ�(1955-)���У����ϳ����ˣ����ڣ���ʿ����ʦ�������ؽ�����Ⱦˮ����������������о����绰��0731-8649208��E-mail: liuyunguo2005@yahoo.com.cn
