
��в������½������Ӧ��FTIR����
Ѧ���������������㣬�ܣ������
(���ϴ�ѧ ұ���ѧ�빤��ѧԺ������ ��ɳ��410083)
ժҪ�����·��ֵ�������ֲ����½Ϊʵ����ϣ��о��ڲ�ͬMn2+Ũ����(0.005��0.2��0.5��1.0��2.0��5.0��10.0 mmol/L)��½��ͬ��֯���ٵĸ���Ҷ�������(FTIR)ͼ�ױ仯���о��������������֯��3 420��2 926 cm-1��������������½�����ӳ����Ͱ�������л����ڵ�Mn2+������������Ϊ���Ե������ʳ���, �京�����ߣ�������Mn2+Ũ�ȵ�����, ��ϳɺ���������, �л��ﺬ�����½�������֯��2 926 cm-1�����ȼ�������ǿ, ˵����½���ڵ��л��������Mn2+, �������O��H���٣�����Mn2+Ũ�ȵ�����, ������������������Ҷ��֯��1 640 cm-1�����������շ��������½Ҷ�а����ᡢ���ĺ͵����������ʺ��������йأ�����������͵������ι����йأ���������˴���ֵ���½��������Ǻϣ���½��������Ҷ��1 060 cm-1�������Ũ���·�ֵ�ı仯��Ҫ������Ĥ֬�����������������½���������йء�
�ؼ��ʣ�
����Ҷ�任�����������½�� Mn2+в������ѧ�����������
��ͼ����ţ�O657.3 ���ױ�־�룺A ���±�ţ�1672-7207(2011)07-1852-06
Physiological response of Phytolacca acinosa to
manganese stress by FTIR spectroscopy
XUE Sheng-guo, WANG Jun, LIU Heng, LEI Jie, LIU Feng-hao
(School of Metallurgical Science and Engineering, Central South University, Changsha 410083, China)
Abstract: A technique based on Fourier transform infrared (FTIR) spectrometry was used to explore the physiological changes in chemical composition of Phytolacca acinosa in a wide range of Mn2+ concentration (0.005, 0.2, 0.5, 1.0, 2.0, 5.0, 10.0 mmol/L). The results show that the absorption bands at 3 420 and 2 926 cm-1 of stem tissues increase firstly and then decrease, indicating the exudation and transporting situation of organic substances which serves as organic osmotic contents to enhance manganese tolerance during low treatments. Meanwhile, the bands at 2 926 cm-1(roots) and 1 640 cm-1(leaves) have different tendencies in accordance with Mn2+ treatments, which shows that the capability to chelate Mn decreases under higher manganese concentration. In addition, the changes of the bands at 1 060 cm-1 differ from each other, suggesting that under the conditions of Mn2+ concentration, the ex-oxidation of membrane lipid increases. It is practical to apply FTIR to compare the chemical differences of the tolerate plants during metal treatments.
Key words: FTIR; Phytolacca acinosa roxb.; manganese stress; chemical composition; tolerance
�ؽ������в��ɽ����Ժͳ־��ԣ�ͨ��ʳ���������ڶ�ֲ�����ڣ�����������彡������������в���������ǻ�����ʶ����ǿ������ұ���մɡ���������ء��������ϡ����ͷ�������ũҩ�ȷ���㷺ʹ���Լ����̿�������������ر�ˮ�͵���ˮ��Mn2+��Ⱦ����õ���ע[1-3]�����г��������Ե�����ֲ�����������ˮ�廷���еĽ������������Ⱦ-ֲ��������(Phytoremediation)����DZ�ڵĸ�Ч�����ۼ��价���Ѻ��ԣ�ͨ����ֲ�ո�����ֲ�����Ч�����������е��ؽ�����Ⱦ[4]��ֲ����ؽ��������Կ�ͨ�������ų�(Metal exclusion)�ͽ�������(Metal accumulation)��2��;�����С������ų���ָ�ؽ�����ֲ�����պ����ų����⣬�����ؽ�����ֲ�����ڵ������ܵ��谭������������ָ�ؽ�����ֲ�������Բ�����������ԵĽⶾ��̬����[5]��������ֲ����һ�ּ��˵Ľ���������ֲ����䳬Ѱ�����ؽ����������������㷺Ӧ������Ⱦ�������о�[6-10]���о���ͬMn2+������������½��ͬ��֯���ٻ�ѧ����ϵIJ��콫�����ڽ�ʾ��Mn2+���Ի�������������з�ӳ����ֲ���л�ϳɷֵĵ��ӣ����ֻ�ѧ�ɷ�ֻҪ�ʺ�������ȶ�����Ʒ�Ĵ���������ͳһҪ����У�����������������ȶ��ġ��������[11]���ø���Ҷ�任�������(FTIR)�о�������½���̶�������Ӧ���ڴˣ�������������FTIR����½��ͬ��֯���ٵĻ�ѧ��ɽ����о�������ΪMn2+��Ⱦ��������Mn2+���Ի����о��ṩ�ο���
1 �����뷽��
1.1 ���鷽��
����½���Ӳ���ʪɳ����ѿ��������0.25 HoaglandӪ��Һ��0.50 HoaglandӪ��ҺԤ����15 d��ѡ������һ�µ���½����������ڲ��������������Mn2+Ũ���趨Ϊ7�֣��ֱ�Ϊ0.005��0.2��0.5��1.0��2.0��5.0��10.0 mmol/L��Mn��MnCl2����ʽ���룬ÿ��ʵ���ظ�3�Ρ�ÿ4 d��1��Ӫ��Һ����������ͨ����ֲ����35 d���ջ���ȥ����ˮϴ�������������ֱ�ȡ��������Ҷ3������105 ��ɱ��30 min��Ȼ����75 ������и���48 h���ò���ַ�������飬��75 ��mɸ��
1.2 ����Ҷ���������
�����������IJ�������(����Χ Ϊ500~4 000 cm-1���ֱ���Ϊ4 cm-1��ɨ���ۼӴ���Ϊ32��)����������Nicolet��˾�����Nexus 670����Ҷ�任��������Ƕ���½��������Ҷ�ķ�ĩ��Ʒ���вⶨ��Ӧ��OMNI������ֱ�Ӳⶨ������ף�������OMNIVE. S.P 5.1ͬ������������ͼ���з�����
2 ���������
2.1 ����FTIR����
��ͬMn2+Ũ������½��ϵ�ĸ���Ҷ�������ͼ1����½��ϵ���������ͼ2����ͼ1�ɼ���3 420 cm-1�������ķ��Ƿ��Ӽ����O��H�����ǻ��������壬��Ҫ��������ά�ء�����ά�ء����ǵ�̼ˮ������[12]������Mn2+Ũ�ȵ����ߣ����շ������Ƶ3 390 cm-1��ͬʱ���α�խ����ͼ2�ɼ�����Mn2+Ũ�ȵ���1.0 mmol/Lʱ������Mn2+Ũ�ȵ����ߣ���������������½���˵����Ũ��Mn2+�ٽ�����½����̼ˮ������ķ��ڣ�ͬʱ������Mn2+Ũ�ȵ����ߣ������Ƥϸ���ڵ��ǻ��������Mn2+�γ��ȶ��Ļ����ʹϸ�������������٣���Mn2+Ũ�ȴ���2.0 mmol/Lʱ����ֵ�ٴγ��������ı仯���ƣ�˵����Ũ��Mn2+�����ƻ��˸����Ƥϸ���ڵ��ǻ��������Mn2+�Ļ��ƣ����¸����Ƥϸ���ڵ��ǻ����������Mn2+�������ֵ���ߡ�

ͼ1 ��ͬMn2+Ũ����������½��ϵ�ĸ���Ҷ�������ͼ
Fig.1 Absorption FTIR spectra in roots of P. acinosa in different Mn2+ concentrations
2 926 cm-1���ҵ����շ��DZ���C��H�������������շ壬��Ҫ������ϸ�����е����ʡ���ά�غ�������֯�ɷ�(ͼ1)����Mn2+Ũ��С��1.0 mmol/Lʱ������Mn2+Ũ�ȵ����ӣ������������(ͼ2)����½�ĸ����������Ի��Ʋ��Ϸ����л�������Mn2+�����ǣ�ǰ��Mn2+����Ũ�Ƚϵͣ�����Mn2+���л�����������С�ڸ������л���������ʣ���������Ƶ������������Mn2+Ũ�ȵ����ӣ�����Mn2+���л����������ʴ��ڸ������л���������ʣ�����Ƶ���ֿ�ʼ�½�����Mn2+Ũ�ȴ���1.0 mmol/Lʱ������Mn2+�����Ľ�һ�����أ������������������������Ƶ�� ������

ͼ2 ��ͬMn2+Ũ������½��ϵ��������仯ͼ
Fig.2 Band height changes in roots of P. acinosa in different Mn2+ concentrations
1 735~1 720 cm-1���ķ�Ϊ֬����ͪ��������ʻ���C=O�����塣��1 735 cm-1����������Mn2+Ũ�ȵ����ߣ�ǰ�ڷ�ֵ���ͣ������Ũ�ȴ����£���ʼ�����������շ壬��ֵ�ﵽ���(ͼ2)�����������Ϊ������½Ĥ�ʹ�������ʹ֬�����ʼ��٣�����֬����ͪ��������ʻ��IJ���Ҳ�����࣬������ڿ�ʼ���ߡ�
1 380 cm-1���������մ��Ǻ���֬������(����Ĥ�Ͱ���)����֯�м������մ�����Mn2+Ũ��Ϊ0.2 mmol/Lʱ����ֵС�������������������ڵ�Ũ��Mn2+�ٽ�����½��������֬������ķ��ڡ����ǣ���Mn2+Ũ�ȴ���0.2 mmol/Lʱ������Mn2+Ũ�ȵĽ�һ���������ֵ�����ȼ�С������ ����Ҳ���ֶ���̲�λ��(��ͼ1)����Ҳ˵������Mn2+Ũ�ȵ����ߣ�ϸ����ͨ�������������̶ȣ�ʹCEC(�����ӽ�������)��ǿ���������ո����Mn2+����ͨ��ϸ�����л���Mn2+����ǿ�����ԣ�����Ũ�ȵ�Mn2+�Ѷ���½������в�ȣ������̶��ֿ�ʼ���ߡ�
1 060 cm-1�������շ�Ϊ̼ˮ������(�����ѻ����������)��C��O���ŵ�������(ͼ1)[12]����Mn2+Ũ��С��0.5 mmol/Lʱ������Mn2+Ũ�ȵ����ߣ������������˵����Ũ��Mn2+�ٽ��������ʵĺϳɣ�Ҳ���������ڻ�ѧ�����ǻ���Mn2+���ã�����C��O��ֵ���ӣ���Mn2+Ũ������ʱ����������ͨ���ܵ�һ��Ӱ�죬��ʹ�����������������䵽��������Mn2+Ũ�ȴ���0.5 mmol/Lʱ��Ĥ���������ƶԷ�ֵ�仯���������ã�����Mn2+Ũ�ȵ����ߣ�Ĥ�������̶ȼ��֬����ͪ���������������ڸ������ۣ������ֵ���ߣ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ����Ũ�ȵ�Mn2+�ƻ��˴˻��ƣ����֬����ͪ����������������٣���ֵ�½�[13-15]��
2.2 ����FTIR����
��ͬMn2+Ũ�����̾����ĸ���Ҷ�������������ֱ��ͼ3��ͼ4����ͼ3�ɼ�����Mn2+Ũ��С��1.0 mmol/Lʱ��3 420 cm-1�������շ�ķ�ֵ�����Ա仯��˵����½����̼ˮ������ɷ�û�����Ա仯����Ҳ������Mn2+Ũ�ȴ���1.0 mmol/Lʱ�����ⲿMn2+��в���£���½���е�����������Ӱ��̶Ƚ�С��Ҳ���ֳ���½��Mn2+������������������Mn2+Ũ�ȵ����ߣ��÷���������������˵������Mn2+Ũ�ȵ����ߣ��ٽ����л���ķ��ں����䣬ͨ������������ǿ����Mn2+�Ŀ����ԣ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ��Mn2+�Ķ��ԼӾ磬��ϸ����ϸ���ڵ��ǻ��������Mn2+��ʹϸ�������������٣������ֵ�½���
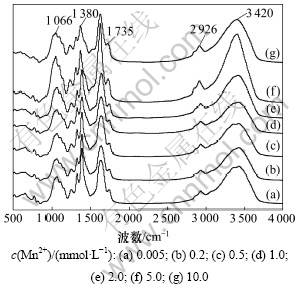
ͼ3 ��ͬMn2+Ũ������½���ĸ���Ҷ�������ͼ
Fig.3 Absorption FTIR spectra in stems of P. acinosa. in different Mn2+ concentrations

ͼ4 ��ͬMn2+Ũ������½����������仯ͼ
Fig.4 Band height changes in stems of P. acinosa in different Mn2+ concentrations
��ͼ3���Կ�����2 926 cm-1�������շ����δ��������λ�ƣ����仯��������Mn2+Ũ�ȵ����ӣ������ǰ�ڱ仯����Mn2+Ũ��Ϊ2.0 mmol/Lʱ�������������½�(ͼ4)����˵����Mn2+Ũ�ȵ���2.0 mmol/Lʱ����Ũ��Mn2+����½�������书��Ӱ�첻����5.0 mmol/Lʱ�������ٽ���½����̼ˮ�������������ǿ��Mn2+���ԣ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ����Ӱ��̼ˮ������ȵĺϳɺ����䡣
��ͼ4���Կ���������Mn2+Ũ�ȵ����ߣ�ǰ�� 1 735~1 720 cm-1�������շ��ֵ�����Ա仯������Mn2+Ũ��Ϊ5.0 mmol/Lʱ�������������շ壬��ֵ�ﵽ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ����ֵ�½������������Ϊ��һ��Mn2+Ũ�ȵĴ̼��»����֬����ͪ���������ʣ�ͨ������������ǿ�����ԣ�������Mn2+���ļӾ磬֬����ͪ���������ʺϳɼ��٣���ֵ�½���
����Mn2+Ũ�ȵ����ߣ�1 380 cm-1�������շ�ķ�ֵ���½����������ڵ�Mn2+Ũ��Ϊ10.0 mmol/Lʱ����ֵ�ٴγ����½�(ͼ4)�����������Ϊ����Mn2+Ũ�ȵ����ߣ�ϸ����ͨ�������������̶ȣ�ʹCEC(�����ӽ�������)��ǿ���������ո����Mn2+����ͨ��ϸ�������ۻ�Mn2+����ǿ�����ԣ�����Ũ�ȵ�Mn2+�Ѷ���½������в�ȣ������̶��ֿ�ʼ���ߣ�������Mn2+Ũ��Ϊ10.0 mmol/L����ֵ�½���ԭ���д���һ���о���
��ͼ3����������Mn2+Ũ�ȵ����ߣ�1 060 cm-1����������մ��ɶ��յ�1 060 cm-1λ�Ƶ�1 052 cm-1�����������α仯�����仯�����ԣ���Mn2+Ũ��С��1.0 mmol/Lʱ������Mn2+Ũ�ȵ����ߣ�����ȱ仯������Mn2+Ũ��Ϊ5.0 mmol/Lʱ�������������շ壬��ֵ�ﵽ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ����ֵ�½�����Mn2+Ũ��Ϊ2.0~5.0 mmol/Lʱ��Ĥ���������ƶԷ�ֵ�仯���������ã�����Mn2+Ũ�ȵ����ߣ�Ĥ�������̶ȼ��֬����ͪ���������������ڸ������ۣ������ֵ���ߡ�����Mn2+Ũ��Ϊ10.0 mmol/Lʱ����Ũ�ȵ�Mn2+�����ƻ��˴˻��ƣ����֬����ͪ����������������٣������ֵ�½�[13-15]��
2.3 Ҷ��FTIR����
Mn2+Ũ�Ȳ�ͬʱ��½Ҷ�ĺ������������ֱ��ͼ5��ͼ6����ͼ5�ɼ�����Mn2+Ũ�ȵ���1.0 mmol/Lʱ������Mn2+Ũ�ȵ����ߣ�3 420 cm-1�������շ�ķ�ֵ�½���˵��ҶƬ��Ƥϸ���ڵ��ǻ��������Mn2+��ʹϸ��������������(ͼ6)������Mn2+Ũ�ȴﵽ2.0 mmol/Lʱ�������������˵������Mn2+Ũ�ȵ����ߣ��ٽ����л���ķ��ں����䣬ͨ������������ǿ����Mn2+�Ŀ����ԣ�ͬʱ��Ũ�ȵ�Mn2+�谭����Щ�л���ĺϳɺ����䡣
2 926 cm-1�����շ�Ϊ����O��H���C��H�����������ص�����Ҫ������ά���غ���Ĥ��ϸ���ڵ���֯�ɷֵ�[12](ͼ5)���������书���йء��ô���ķ�ֵ����Mn2+Ũ�����������ߺ��½�(ͼ6)������˵��ǰ����½���ڵ��л��������Mn 2+���������O��H �ļ��٣�����Mn2+�����ļ��أ�������������������
�ڵ���ԴMn�����£�1 650~1 620 cm-1���ķ�ֵ�ڴ���ǰ���½���˵������Mn2+Ũ�ȵ����ߣ��������ṹ�У��ļ�������Ľ��������Mn2+Ũ�ȵ����߶�����(ͼ6)������Mn2+Ũ��Ϊ2.0 mmol/Lʱ�������������շ壬���������ڲ������ӵ�Mn2+�յ��������ᵰ�ס�������ص����ʰ��ᵰ��һЩ���ϳɣ�Didierjean��[16]����Ϊ��Щ�ؽ���в���յ������ܾ��б���ֲ��ϸ�������ؽ������������ã����ֵ��������շ��������½Ҷ�а����ᡢ���ĺ͵����������ʺ��������йء���Mn2+Ũ��Ϊ5.0��10.0 mmol/Lʱ��������½��������Ǹ�Ũ�ȵ�Mn2+���ƻ����ؽ���в���յ����ϳɻ������¡�

ͼ5 ��ͬMn2+Ũ������½Ҷ�ĸ���Ҷ�������ͼ
Fig.5 Absorption FTIR spectra in leaves of P. acinosa in different Mn2+ concentrations
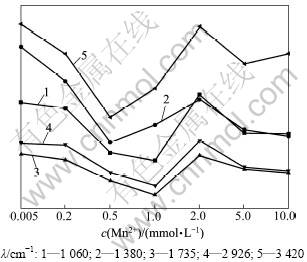
ͼ6 ��ͬMn2+Ũ������½Ҷ��������仯ͼ
Fig.6 Band height changes in leaves of P. acinosa in different Mn2+ concentrations
����Mn2+Ũ�ȵ����ߣ�1 380 cm-1�����ķ�ֵ�����Ƚ����������ƣ�����Mn2+Ũ��Ϊ2.0 mmol/Lʱ���ֳ������½�����˵������Mn2+Ũ�ȵ�����ϸ����ͨ�������������̶ȣ��������ո����Mn2+����ͨ��ϸ�����л���Mn2+����ǿ�����ԣ�����Ũ�ȵ�Mn2+����½������в�ȣ������̶��ֿ�ʼ���ߣ���Mn2+Ũ��Ϊ2.0 mmol/Lʱ���ֳ������½���ԭ������Ǹ�Ũ�ȵ�Mn2+������ϸ���ں���֬�����������
��ͼ5�ɼ�����Mn2+Ũ��С��1.0 mmol/Lʱ������Mn2+Ũ�ȵ����ߣ�1 060 cm-1�������շ�ķ�ֵ�½���˵����Mn2+Ũ������ʱ���������ʵ������ܵ�Ӱ�죬��ֵ���ͣ���Mn2+Ũ�ȴ���1.0 mmol/Lʱ��ֵ������������Ĥ�������̶ȼ��֬����ͪ����������������Ҷ�����ۣ������ֵ���ߣ�����Mn2+Ũ��Ϊ5.0��10.0 mmol/Lʱ����Ũ�ȵ�Mn2+�ƻ��˴˻��ƣ����֬����ͪ����������������٣���ֵ�½�[13-15]��
3 ����
(1) ����Ͱ�������л��������ڵ�Mn2+����������������Ϊ���Ե������ʳ��֣��京�����ߣ�����Mn2+�����ļ��أ���ϳɺ����䶼�����ޣ�������Ȼ�½��������䵼��֯�б������ԡ���Mn2+Ũ��Ϊ0.2 mmol/Lʱ������2 926 cm-1���ķ�ֵ�ȼ�������ǿ, ˵����½���ڵ��л��������Mn2+���������O��H���٣�����Mn2+�����ļ��أ�����������������������Mn2+Ũ��Ϊ2.0 mmol/Lʱ��Ҷ��1 640 cm-1�����������շ��������½Ҷ�а����ᡢ���ĺ͵����������ʺ��������йأ�����������͵������ι����йأ���������˴���ֵ���½������Ǻϡ�
(2) ��½��Mn2+�������Դ���һ���ٽ�ֵ�����������ٽ�ֵʱ��ֲ��ͬ�����ܵ���������Ҫ������Ĥ֬����������������Ҷ��1 060 cm-1����ֵ�ı仯��֤������һ�㡣
(3) ��½��������Ҷ��֯�����ࡢ��������л�������ʺ�����Mn2+Ũ�����߶��仯�ϴ�����֯�����ʺ����仯��������Mn2+Ũ��С��2.0 mmol/L���ⲿMn2+в���£���½����֯�е�����������Ӱ���С���л���ĺϳɺ��������û���ܵ�Ӱ�죬Ҳ������½���н�ǿ��Mn2+����������
�ο����ף�
[1] �̹�ƽ, ����ϼ, ������. ��������������ҵ��Ⱦ����������[J]. �й��������, 2003, 19(4): 56-59.
CAI Gu-ping, GE Xiao-xia, ZENG Guang-ming. Investigation and evaluation of the pollution of manganese sulphate in Huangxing Town[J]. Environmental Monitoring in China, 2003, 19(4): 56-59.
[2] Aydinalp C, Marinova S. Distribution and forms of heavy metals in some agricultural soils[J]. Polish Journal of Environmental Studies, 2003, 12(5): 629-633.
[3] ����˳, ����ǿ, ��С��, ��. ���ݰ�����������-ˮ�����������������Ԫ����Ǩ�Ƹ�����Ӱ��[J]. ��ѧͨ��, 2003, 48(19): 2073-2078.
WANG Fu-shun, LIU Cong-qiang, LIANG Xiao-bing, et al. Microbiological activity across the sediment-water interface of Lake Aha and its influence upon translocation and accumulation of trace elements[J]. Chinese Science Bulletin, 2003, 48(19): 2073-2078.
[4] Myrna E W. Phytoremeditaion on the brink of commercialization[J]. Environment Science and Technology, 1997, 31(4): 182-186.
[5] Baker A J M, Brooks R R. Terrestrial higher plants which hyperaccumulate metallic elements: A review of their distribution, ecology and phytochemistry[J]. Biorecovery, 1989, 1(2): 81-96.
[6] Ѧ����, ��Ӣ��, ����, ��. �й��״η��ֵ��̳�����ֲ��: ��½[J]. ��̬ѧ��, 2003, 23(5): 935-937.
XUE Sheng-guo, CHEN Ying-xu, LIN Qi, et al. Phytolacca acinosa Roxb. (Phytolaccaceae): A new manganese hyperaccumulator plant from Southern China[J]. Acta Ecological Sinica, 2003, 23(5): 935-937.
[7] XUE Sheng-guo, CHEN Ying-xu, Reeves R D, et al. Manganese uptake and accumulation by the��hyperaccumulator plant Phytolacca americana Roxb. (Phytolaccaceae)[J]. Environmental Pollution, 2004, 131(3): 393-399.
[8] XUE Sheng-guo, CHEN Ying-xu, Baker A J M, et al. Manganese uptake and accumulation by two populations of Phytolacca acinosa Roxb. (Phytolaccaceae)[J]. Water Air and Soil Pollution, 2005, 160(1/3): 3-14.
[9] XU Xiang-hua, SHI Ji-yan, CHEN Ying-xu, et al. An investigation of cellular distribution of manganese in hyperaccumlator plant Phytolacca acinosa Roxb. using SRXRF analysis[J]. Journal of Environmental Sciences-China, 2006, 18(4): 746-751.
[10] Ѧ����, Ҷ��, �ܷ�, ��. �̳�����ֲ�ﴹ����½���϶�[J]. ��̬ѧ��, 2008, 28(12): 6344-7347.
XUE Sheng-guo, YE Sheng, ZHOU Fei, et al. Phytolacca americana L.(Phytolaccaceae), Pokewood: A manganese hyperaccumulator plant[J]. Acta Ecologica Sinica, 2008, 28(12): 6344-6347.
[11] ������, �����, ����, ��. ������½���̶�������Ӧ��FTIR�о�[J]. ����ѧ�������, 2008, 28(3): 582-585.
REN Li-min, CHENG Ze-feng, LIU Peng, et al. Studies on the physiological response of Phytolacca americana to manganese toxicity by FTIR spectroscopy[J]. Spectroscopy and Spectral Analysis, 2008, 28(3): 582-585.
[12] ½����, Ԭ�鸣, ���ͨ, ��. �ִ����������������[M]. ����: �й�ʯ��������, 2000: 19-26.
LU Wan-zhen, YUAN Hong-fu, XU Guang-tong, et al. NIR spectra analysis[M]. Beijing: China Petrochemical Press, 2000: 19-26.
[13] �ٹ���, ���»�, ���غ�, ��. Cu��Ⱦ��С������������ع�ϵ�о�[J]. ����ѧ�������, 2006, 26(7): 1272-1276.
CHI Guang-yu, LIU Xin-hui, LIU Su-hong, et al. Studies of relationships between Cu pollution and spectral characteristics of TritiZnm aestivum L[J]. Spectroscopy and Spectral Analysis, 2006, 26(7): 1272-1276.
[14] ��˼��, ���»�, ���, ��. �ؽ���пв�ȵİײ�ҶƬ������Ӧ�о�[J]. ����ѧ�������, 2007, 27(9): 1797-1810.
CHEN Si-ning, LIU Xin-hui, HOU Juan, et al. Study on the spectrum response of Brassica campestris L leaf to the zinc pollution[J]. Spectroscopy and Spectral Analysis, 2007, 27(9): 1797-1810.
[15] ������, ����, ���, ��. ���յ�ֲ���ϸ���ڻ�ѧ�ɷֱ仯��FTIR����[J]. ����ѧ�������, 2008, 28(5): 1067-1070.
ZHANG Xiao-bin, LIU Peng, LI Dan-ting, et al. FTIR Spectroscopic characterization of chromium-induced changes in root cell wall of plants[J]. Spectroscopy and Spectral Analysis, 2008, 28(5): 1067-1070.
[16] Didierjean L, Frendo P, Nasser W, et al. Heavy metal responsive genes in maize: Identification and comparison of their expression upon various forms of abiotic stress[J]. Planta, 1996, 199(1): 1-8.
�ո����ڣ�2010-10-15�������ڣ�2010-12-28
������Ŀ�����ҹ�����(����)��ҵ������Ŀ(200909065��201109056)��������Ȼ��ѧ����������Ŀ(40771181)���й���ʿ���ѧ����������Ŀ(20080430565)���й���ʿ���ѧ�����ر�������Ŀ(200801119)
ͨ�����ߣ�Ѧ����(1970-)���У����Ͻ����ˣ���ʿ�������ڣ�������Ⱦ�������������ɽ��������̬�ָ������о����绰��13787148441��E-mail: sgxue70@yahoo.com.cn
