
��������������˾�ATCC23270���ʵ���
ѡ������ȡ���������
�Ľ������� ������ ٻ�����������ųɹ�ŷ���������
(���ϴ�ѧ ��Դ�ӹ������﹤��ѧԺ ����ұ��������ص�ʵ���ң����� ��ɳ��410083)
ժ Ҫ��
ժ Ҫ��ͨ��������ȡʱ�䡢����Һ�����Һ��Ħ�����Լ��ܵ���Һ�����ϸ����ƻ��������ø�ıȻ���(�����ϸ������������)��Ӱ�죬����ѡ������ȡAcidithiobacillus ferrooxidans ATCC23270���ʵ������ݿ�ʵ�鷽���������羵��˫���Ӿʵ�鷽��������������ǰ��ϸ������̬�Ͳ�ͬ�����ڵ�ϸ�����ʿռ䵰�ױ�����졣�о�������������ʵ����ݿ�ʵ������Ϊ��ϸ������Ϊ0.1 g(ʪ��)����Һ�������Ϊ10 mL����Һ������ʱ��Ϊ15 min, ����Һ�����Һ��Ħ����Ϊ1?1���ڸ��������£�ϸ�����������ʿռ��С���ڵ���Һ�����£�ϸ�����ʿռ����ͣ���Ĥ�����ƣ����ʿռ��ڵ����ͷų�����˫���Ӿ��ʾ���ʵ�����������С��Է�����������Ϊ�����ڲ�ͬ������ʱ���ڵ����ʵ��ױ�����ڲ��졣
�ؼ��ʣ�
Acidithiobacillus ferrooxidans�����ʵ�����˫���Ӿ��ƻ��������ø���������ݿ˷���
��ͼ����ţ�Q93 ���ױ�ʶ�룺A ���±�ţ�1672-7207(2009)04-0845-06
Selective extraction and differential electrophoregrams analysis of periplasmic proteins of Acidithiobacillus ferrooxidans ATCC23270
XIA Jin-lan, WANG Jing, ZHANG Qian, ZHANG Rui-yong, ZHANG Cheng-gui,
OUYANG Xu-dong, QIU Guan-zhou
(Key Laboratory of Biometallurgy of Ministry of Education, School of Minerals Processing and Bioengineering,
Central South University, Changsha 410083, China)
Abstract��The selective extraction of periplasmic proteins by osmotic shock was investigated. The effects of the extraction efficiency and the pollution from the cytoplasmic proteins were tested and monitored, respectively, in terms of the specific activity of malate dehydrogenase (MDH). And the morphology and differential expression of the periplasmic proteins of Acidithiobacillus ferrooxidans were analyzed by using TEM and 2-D electrophoresis. The results show that the preferable schema of osmotic shock is founded as follows: the wet mass of cell is 0.1 g, the molar ratios of hypertonic saline to hypotonic saline is 1?1, the volume of the osmotic solvent is 10 mL and the extracting time is 15 min. The TEM images show that under the treatment from hypertonic to hypotonic environment, the cell outer membrane is selectively burst, which makes the periplasmic space protein released. The 2-D electrophoregrams display the most of periplasmic proteins are small molecular weight protein and cells growth in different phase present difference in periplasmic proteins expression.
Key words: Acidithiobacillus ferrooxidans; periplasmic proteins; two-dimensional electrophoresis; MDH activity; osmotic shock extraction
��������������˾�(Acidithiobacillus ferrooxidans)��һ�������Ի�����������������ϸ��������ͨ�������������������ʻ���������������[1]��A. ferrooxidans����Դ���ʵ��������ù���ʮ�ָ��ӣ�Ȼ�����ù�����Ҫ������ϸ�����ʿռ� ��[2]��������;����Ҳ�кܶ�Ԥ�⵰������˫����ø����øQ������ԭø��������ø������-������������ԭø�ȴ��������ʿռ�[3-6]���������Ƕ�A. ferrooxidans�������������̵��о����٣�Ŀǰ������ȷ������Ԫ����ĵ������û��ơ���Ȼ�����о��߶Բ�ͬ��Դ����A. ferrooxidans�ĵ�������ѧ������һ���̶ȵ��о��ͷ���[7-8]�����ǣ�����ȫ���������Ӵ������죬���о����о���������˺ܴ����ѡ���������о��ߴ��ػ�ϸ���ռ����֣���ϸ�����ʿռ�Ⱦֲ�������˫���Ӿ�������[9]���ػ��ռ䵰������ܵ����������٣����ҹ���������Ӷ��������о��������Ѷȡ��ڴˣ��������߶����ʵ�����ȡ�����������ݿ˷������Ż������Ե������������в�ͬ����ʱ�ڵ�A. ferrooxidans�������ʵ�����˫���Ӿ�������ͼ�Աȷ������Ա�Ϊ���õ���Ӿ��������ɸѡA. ferrooxidansϸ����������������عؼ����춨������
1 �����뷽��
1.1 ���ꡢ����������������
A. ferrooxidans ATCC 23270��Դ������ģʽ�����ռ����ġ�ʹ��9K����������Һ���������Ծ��ֽ��л�ʹ������������ɷ�Ϊ��(NH4)2SO4��3.0 g/L�� MgSO4��7H2O��0.5 g/L��KCl��0.1 g/L��K2HPO4��0.5 g/L�� Ca(NO3)2��0.01 g/L��S��5 g/L����5 mol/L��H2SO4����������ʼpHֵ����Ϊ2��ϸ����Ԥ�������5 L����ƿ��30 ��������У�������Ϊ10%��Ũ��Ϊ 108 ��/mL��ͨ������������������������Ѫ����������[10]����ʱ��Ծ�����ͼ�ⶨ��������������ϸ���������ߡ�
1.2 ����Һ����Ҫ�Լ�
������Һ���Ϊ��10 mmol/L��Tris(pH8.0)����Һ��20%���ǣ�25 mmol/L EDTANa2��������ҺΪ 5 mmol/L MgCl2��ƻ��������ø�ⶨ�Լ��й����Ͼ��������﹫˾����100 mg����˹����G-250���� 50 mL 90%�Ҵ��У�����85%������100 mL���������ˮ��1 L���ڳ��±��棬�Ƶÿ���˹����G-250���ײⶨ�Լ������Ỻ��Һ(PBS)�����Ϊ��NaCl��8.0 g��KCl��0.2 g��Na2HPO4��12H2O��3.48 g��KH2PO4��0.2 g��������ˮ��1 L����ȡ1 mgţѪ�嵰�ף�����1 mL����ˮ�У��Ƴ�1 mg/mL�����ʱ�Һ��
����pH�ݶȸɽ���(IPG������pHֵΪ3~10������Ϊ24 cm)��IPG����Һ(pHֵΪ3~10)ΪAmersham Pharmacia��˾��Ʒ����������Է�����������ΪMBI��˾��Ʒ��
˫���Ӿˮ��Һ��8 mol/L���أ�2 mol/L���壬2% CHAPS��18 mmol/L DTT��0.5 mmol/L PMSF��0.5% IPG buffer��0.002%�������ɡ�
ƽ���Һ��0.05 mol/L Tris����Һ��6 mol/L���أ�30%(�������)���ͺ�2%(��������)SDS��ɣ�pH=8.8��
1.3 ���ʵ�����ȡ�����Ż�
1.3.1 ǰ�ڴ���
�ö�����ֽ���˴��ڶ��������ڵ�A. ferrooxidansϸ������������Һ�������ռ�ϸ��������10 000 r/minʱ��4 ������15 min�վ�����������9K������������(����������)ϴ3�Σ�����pHֵΪ2.0��H2SO4��Һϴ��3�Ρ�ƽ����Ϊ���ɷݱ��ã�ÿ�ݴ�������ϸ��ʪ��ԼΪ0.1 g��
1.3.2 ���ʵ���ԭʼ��ȡ����
ȡ1���ռ��ľ����ڼ������Һ1 mL�������»�������ҡ��30 min���ڵ�������(6 000 r/min)�����ռ�������ټ������Һ1 mL�����ȣ���ԡ30 min����4 �桢ת��Ϊ16 000 r/min����10 min���ռ����壬����ȡ���ʵ�����Һ[11]��
1.3.3 �����Ż�
ͨ������Һ����ʱ�䣬��Һ����������Լ��������Һ�����Һ�ı����ĸı�۲���ȡЧ�ʱ仯�������������Bradford���ⶨ������Ũ��[12]������Ũ�Ⱥ����������ܵ�������
1.3.4 �����ȡ������ȷ��
�����ʵ�����ȡҺ�а��ʵ�����Ⱦ����Ϊԭ���ݲ���������ѭ��øƻ��������ø�Ļ�������ȡ�ܵ�����֮��ȷ�������ȡ������ƻ��������ø(Malate dehydrogenase, MDH)��һ�ֶ�����ø���ף��㷺���������ϸ����ԭ��ϸ���У���������ԭ��лø�е���Ҫ��Ա֮һ����ϸ���ʿռ�ı�־ø֮һ[13]��ͨ�����MDH���Կ��Ե�֪ϸ���ʵ�����Ⱦ�����MHD����Խ�ߣ�˵����ȾԽ���ء�MDH�����ܹ���ƻ���������ɲ������ᣬ�����ܹ����������ỹ ԭ[14]����MDH�����£�ƻ���ᱻ���������ɲ������ᣬNAD+������ø�ĸ�ø����NAD+��Ϊ��������������ΪNADH+H+��NADH��340 nm�������շ壬NAD+û�����շ壬��ˣ�ͨ�����340 nm������ȵı仯���ԲⶨMDH����[15]��MDH���Լ�������������ø���Լ��㷽���ο��Ͼ��������﹫˾��Ʒƻ��������ø�ⶨ�Լ���˵���顣
1.4 ��ͬ����ʱ�ڵ�A. ferrooxidans���ʵ���˫���Ӿͼ�ײ���Ƚ�
���������ȡ�����ֱ�Ե�����������������A. ferrooxidans�����ڣ��ȶ��ں�˥����ϸ����ȡ���ʵ��ף������䶳����ɷ�ĩ״����˫���Ӿ����ǰ��ˮ��Һ�ܽ��ͨ��Bradford���ⶨŨ�ȣ���ÿ������300 ?g���е���˫���Ӿ[16]������pH�ݶȸɽ���pHֵΪ3~10��30 Vˮ��12 h����ѹ��8 kV���۽�5 h������Сʱ��ԼΪ80 000 V?h��ƽ����IPG��ˮƽ����12%��SDS-PAGE���ϣ���0.8%���۵���֬�Ƿ⽺���̶�IPG������еڶ����Ӿ�������ÿ���˹����Ⱦɫ��Ⱦɫ��
2 ���������
2.1 ���ʵ��������ȡ������ȷ��
�����ʵ�����ȡ�����������������ǰ���£����ı���Һ������ʱ�䣬�����ܵ�������ƻ��������ø���ԡ�������ͨ��Bradford���ⶨŨ�Ⱥ�����������õ���ƻ��������ø��������ƻ��������ø�ⶨ�Լ��м�⣬������1��ʾ���ӱ�1�ɼ�����������ʱ������ӣ����ʵ�����ȡ��Ҳ��Ӧ������ȾҲԽ��Խ���أ�������ʱ������(15 min)��Ϊƻ��������ø�Ȼ�����Сֵ��˵����ʱ���ʵ�����Ⱦ���١�
��1 ��Һ����ʱ���ƻ��������ø�Ȼ��Ե�Ӱ��
Table 1 Effects of extraction time on specific activity of MDH

������ʱ��Ϊ15 min�����ʵ�����ȡ����������������ʱ���ı�������Һ�����Һ��Ħ���ȣ������ܵ�������ƻ��������ø���ԣ�ʵ�����ֵ��������2��ʾ����2�������ڸ���ҺĦ���������ǰ���£�ͨ�����ӵ���Һ������������ҺĦ�������õ������ʵ�������Ӧ���ӣ�ƻ��������ø�Ȼ����ȼ��ٺ����ӣ��ڸ���Һ�����Һ��Ħ����Ϊ1?1ʱ������Сֵ��˵���ڸ�Ħ�����°��ʵ��������Ⱦ���٣�Ч����á�
��2 ����Һ�����ҺĦ���ȶ�ƻ��������ø�Ȼ��Ե�Ӱ��
Table 2 Effects of molar ratios of hyper- to hypo-osmotic pressure on specific activity of MDH

������ʱ��Ϊ15 min������Һ�����Һ��Ħ����Ϊ1?1��ǰ���£��ı�������Һ�����Һ������������쵰������ȡ����ƻ��������ø�ıȻ��Եı仯��ʵ�����ֵ�ͽ�����3��ʾ����3������������Һ����������ӣ����ʵ�����ȡ������Ӧ����ƻ��������ø�Ȼ����ȼ��ٺ����ӣ��������Ϊ10 mL����С��˵���ڸ������°��ʵ�����Ⱦ��Խ��٣���ȡЧ���Ϻá�
��3 ��Һ�����ƻ��������ø�Ȼ��Ե�Ӱ��
Table 3 Effects of osmotic solvent volume on specific activity of MDH

�ɼ����Ϻ��ʵIJ���ֵΪ���������ʱ��Ϊ15 min������Һ�����Һ�����Ħ����Ϊ1?1����Һ����������Ϊ10 mL���ڴ���������ȡ�������ʵ��������ϸߣ������ʵ�����Ⱦ�̶���С��
�������ȡ������ԭʼ��ȡ�����ֱ���ȡ3�ݾ������ʵ��ף��ⶨŨ�Ⱥ���Ϊ�����������ⶨ������ȡҺ��ƻ��������ø���ԣ�������4��ʾ���ɼ�������������ȡ�ĵ��������ԭʼ������߽�50%��MDH�Ȼ��Խ�����20%���ң���ϸ���ʵ�����Ⱦ����20%�������Ⱦ����1/2���ϣ�˵�������ķ��������ӵ��ʵ�ǰ���¼�������Ⱦ��ȷ��ΪA. ferrooxidans���ʵ�����ȡ���Ż�������
��4 ���������ݿ˷���ԭʼ�����ķ���Ч���Ա�
Table 4 Comparison in extraction effect between modified and unmodified osmotic shock methods

2.2 ���ݿ˷�������A. ferrooxidansϸ����羵����
������羵������A. ferrooxidans ϸ���������ݿ˷�������ϸ�����ͻ�����״���������ͼ1��ʾ���ɼ���ϸ���ڸ����͵����Ļ����µ�ϸ��������̬���в�ͬ���ڸ��������£�ϸ����ˮ������������ϸ��������ϸ����Ĥ(PM)��ϸ����Ĥ(OM)֮��Ŀ�϶(PS)��С�����ʿռ��С����ϸ����������Һ����һ��ʱ��֮��ϸ����̬����һ���仯��ϸ�������ˮӿ��ϸ��ϸ�����ʿռ䣬����ϸ�����ʿռ伱���� �ͣ���ʱ���ʿռ��ڵ�������֮��������������Ĥ���ƻ���
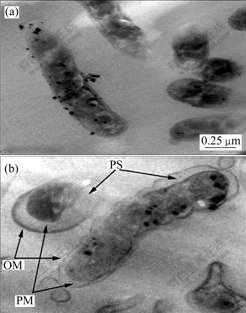
(a) ����Һ��(b) ����Һ
ͼ1 �ֱ��ڸ����͵��������д������A. ferrooxidansϸ��ϸ���ij�����Ƭ��羵ͼ
Fig.1 TEM images of A. ferrooxidans cells after treated by hypertonic solution and hypotonic solution
2.3 ��ͬ������A. ferrooxidans���ʵ���˫���Ӿ�Աȷ���
ͼ2��ʾΪ������Ԫ��������е�A. ferrooxidans�������ߡ��ɼ������Ե�������Ϊ��Դ��9K�������У�A. ferrooxidans�ڽ��ֺ�25~150 h�ڴ��ڶ��������ڣ�150~300 h�ڼ�Ϊ�ȶ��ڣ�300 h�����˥���ڡ���������ѡ������ʵ�����ȡ�������ֱ�Զ��������ڡ��ȶ��ں�˥���ڵ�ϸ�����ʵ�������ѡ������ȡ��Ȼ����˫���Ӿ������ȡ�����ʵ����н�һ�����룬�õ���Ӿ����ͼ3��ʾ�����У���Ϊ1��ԲȦ��Ϊͼ3(a)��3(b)�п��ܵ���ͬ�����ʰߵ㣬��Ϊ2��ԲȦ��Ϊͼ3(b)��3(c)�п��ܵ���ͬ�����ʰߵ㣬��Ϊ3��ԲȦ��Ϊͼ3(a)��3(b)��3(c)�п��ܵ���ͬ�����ʰߵ㣬��Ϊ4��ԲȦ��Ϊ���Բ�ͬ�����쵰���ʰߵ㡣
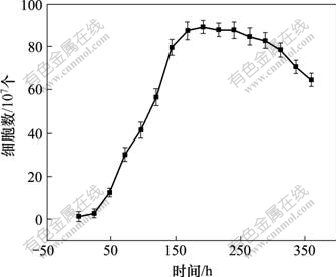
ͼ2 S0��������A. ferrooxidans����������
Fig.2 Growth curve of A. ferrooxidans in elemental sulfur medium

(a) 100 h����������; (b) 200 h�ȶ���; (c) 300 h˥����
ͼ3 ��������������A. ferrooxidans��ͬ�����ڵ����ʵ���˫���Ӿͼ��
Fig.3 2D-electrophoregrams of periplasmic proteins in different growing phases of A. ferrooxidans grown in elemental sulfur media
��ͼ3��֪���ڵ������������У�A. ferrooxidans���ʵ�������Ϊ��
a. ��С��Է�����������Ϊ��������������ʿռ䵰��Ϊ����������ԭ�ĵ���ת�����йأ�С��Է������������ܸ������ڵ��أ�����Ѹ�٣�
b. ��Ϊ���Ե���[11]������������ʿռ�Ϊ���Ի����йأ����Ե��������Ի�����Զ��ȵ�㣬���������ȶ����ڣ�
c. ���������ʵ���(ͼ3(a))����Է����������Ը�ΪƫС�������ߵ��λ�ڼ��Զˣ��ߵ�����������
d. ���������ȣ��ȶ��ڵ����ʵ���(ͼ3(b))������Ȼ�Լ��Ե���Ϊ�������װߵ�Ҳ�������������в������Ƶ��װߵ㣬���ȶ�����һЩ����Է����������ױ�������(ͼ3(b)��ԲȦ2��ʾ)��ͬʱijЩ�ڶ����ڱ���ĵ���(��ͼ3(a)�е�ԲȦ4)���ȶ�����δ�����
e. ������ں��ȶ�����ȣ�˥�������ʵ��ײ��ְߵ����ȶ�������(ͼ3(c)��ԲȦ2��ʾ)�����ߵ�������������(ͼ3(c))���Ҷ�Ϊ���Ե���(ͼ3(c)��ԲȦ4��ʾ)��˵�������Ѿ��в���ϸ���Ѿ�˥���ܽ����ϸ���ʵ�����Ⱦ��ͨ��������Ʒ�е�ƻ��������ø���ԣ�����ø���ԼӴ�(���δ����)��Ҳ֤������㣻
f. 3����ͬʱ��ֻ����һ�鵰�װߵ�ʼ�մ���(��ͼ3��ԲȦ3��ʾ)������Ϊij����Ե��ף���λ�����Զˡ�
3 �� ��
a. A. ferrooxidans ATCC23270���ʵ�����ȡ�ĺ��ʷ���Ϊ������0.1 g(ʪ��)ϸ������10 mL����Һ����15 min������ȥ���壬����10mL����Һ��������ϸ��15min������ȡ����õ����ʵ��ס�
b. ���ʵ�����������С��Է�����������Ϊ����
c. ���������ȶ��ڶ�Ϊ���Ե��ס�
d. ���������ʵ��ķ������ձ�ƫС�������ߵ��λ�ڼ��Զˣ��ߵ�����������
e. �ȶ��ڵ����ʵ��װߵ�Ҳ�������������в��ֵ��װߵ�����������ƣ����ȶ�����һЩ����Է����������ױ������ӣ�ͬʱ��ijЩ�ڶ����ڱ���ĵ������ȶ�����δ�����
f. ˥�������ʵ��ײ��ְߵ����ȶ������ƣ��ߵ��������ӣ��Ҷ�Ϊ���Ե��ס�
g. 3����ͬʱ��ֻ����һ�鵰�װߵ�ʼ�մ��ڣ�����Ϊij����Ե��ף���λ�����Զˡ���Щ���Ϊ��һ���о����ʿռ��ڹ����Ե��춨�˻�����
�ο����ף�
[1] ���, �����, �ƾշ�, ��. ��������������˾��ı��ط���[J]. ���ϴ�ѧѧ��: ��Ȼ��ѧ��, 2006, 37(3): 472-475.
YANG Yu, ZHANG Yan-fei , HUANG Ju-fang, et al. Preservation of Acidithiobacillus ferrooxidans[J]. Journal of Central South University: Science and Technology, 2006, 37(3): 472-475.
[2] Rawlings D E. Characteristics and adaptability of iron- and sulfur- oxidizing microorganisms used for the recovery of metals from minerals and their concentrates[J]. Microbial Cell Factories, 2005, 4: 1-13.
[3] Rohwerder T, Wolfgang S. The sulfane sulfur of persulfides is the actual substrate of the sulfur-oxidizing enzymes from Acidithiobacillus and Acidiphilium spp.[J]. Microbiology, 2003, 149(7): 699-710.
[4] Prange A, Engelhardt H, Tr��per G, et al. The role of the sulfur globule proteins of Allochromatium vinosum: mutagenesis of the sulfur globule protein genes and expression studies by real time RT-PCR[J]. Arch Microbiol, 2004, 182(2): 165-174.
[5] Sugio T, Katagiri T, Inagaki K, et al. Actual substrate for elemental sulfur oxidation by sulfur: ferric ion oxidoreductase purified from Thiobacillus ferrooxidans[J]. Biochim Biophys Acta, 1989, 973: 250-256.
[6] �ųɹ�, �Ľ���, �� ��, ��. ����������ϸ��������ϵͳ�о���չ[J]. ���\��ͨ��, 2007, 13(1): 59-65.
ZHANG Cheng-gui, XIA Jin-lan, WANG Jing, et al. A model of sulfur oxidation system of Acidithiobacillus spp[J]. Biotechnology Information, 2007, 13(1): 59-65.
[7] HE Zhi-guo, HU Yue-hua, ZHONG Hui, et al. Preliminary proteomic analysis of Thiobacillus ferrooxidans growing on elemental sulphur and Fe2+ separately[J]. Journal of Biochemistry and Molecular Biology, 2005, 38(3): 307-313.
[8] Ramirez P, Guiliani N, Valenzuela L, et al. Differential protein expression during growth of Acidithiobacillus ferrooxidans on ferrous iron, sulfur compounds, or metal sulfides[J]. Applied and Environmental Microbiology, 2004, 70(8): 4491-4498.
[9] Chi A, Valenzuela L, Beard S, et al. Periplasmic proteins of the extremophile Acidithiobacillus ferrooxidans: A high throughput proteomic analysis[J]. Mol Cell Proteomics, 2007, 6(12): 2239-225.
[10] �� ƽ, ������, �����. �ؽ����������������̬ЧӦ[J]. Ӧ����̬ѧ��, 1997, 8(2): 218-224.
GONG Ping, SUN Tie-heng, LI Pei-jun. Ecological effect of heavy metals on soil microbes[J]. Chinese Journal of Applied Ecology, 1997, 8(2): 218-224.
[11] Nossal N G, Heppel L A. The release of enzymes by osmotic shock from Escherichia coli in exponential phase[J]. Journal of Biological Chemistry, 1966, 241(13): 3055-3062.
[12] Bradford M. A rapid and sensitive for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding[J]. Analytical Biochemistry, 1976, 72(5): 248-254.
[13] �������� G. ø�������ֲ�[M]. �Ϻ�: �Ϻ���ѧ����������, 1983.
Gilberte G. Enzyme analysis manual[M]. Shanghai: Shanghai scientific and Technical Press, 1983.
[14] �� ��, �� ��. ɳ�ž�CWDMsƻ��������ø�ļ��[J]. �й���̬ѧ��־, 2001, 13(4): 216-217.
JIANG Yan, WANG He. Determination of MDHs of the CWDMs of Salmonella Spp[J]. Chinese Journal of Microecology, 2001,13(4): 216-217.
[15] ���Ǻ�, ��־��, ������, ��. ϸ����NAD(P)Hˮƽ�����������о���չ[J]. ��ѧ��չ, 2004, 16(6): 962-968.
CHEN Ya-hong, LIU Zhi-hong, CAI Ru-xiu, et al. Determination of the Intracellular NAD(P)H Level[J]. Progress in Chemistry, 2004, 16(6): 962-968.
[16] Angelika G, Walter W, Michael D. Current two-dimensional electrophoresis technology for proteomics[J]. Proteomics, 2004, 4(12): 3665-3685 .
�ո����ڣ�2008-10-05�������ڣ�2008-12-25
������Ŀ��������Ȼ��ѧ����������Ŀ(50674101)����973���ƻ���Ŀ(2004CB619201)��������Ȼ�������о�Ⱥ���ѧ����������Ŀ(50621063)
ͨ�����ߣ��Ľ���(1964-)���У����������ˣ���ʿ�����ڣ���������ѧ�����﹤���о����绰��0731-88836944��E-mail: jlxia@mail.csu.edu.cn
[1] ���, �����, �ƾշ�, ��. ��������������˾��ı��ط���[J]. ���ϴ�ѧѧ��: ��Ȼ��ѧ��, 2006, 37(3): 472-475.
[6] �ųɹ�, �Ľ���, �� ��, ��. ����������ϸ��������ϵͳ�о���չ[J]. ���\��ͨ��, 2007, 13(1): 59-65.
[J]. Applied and Environmental Microbiology, 2004, 70(8): 4491-4498.
[10] �� ƽ, ������, �����. �ؽ����������������̬ЧӦ[J]. Ӧ����̬ѧ��, 1997, 8(2): 218-224.
[13] �������� G. ø�������ֲ�[M]. �Ϻ�: �Ϻ���ѧ����������, 1983.
[14] �� ��, �� ��. ɳ�ž�CWDMsƻ��������ø�ļ��[J]. �й���̬ѧ��־, 2001, 13(4): 216-217.
